
乌金山植物群落物种多样性的垂直分布格局
【类型】期刊
【作者】牛常青,曲波,牛霞霞(晋中学院生物科学与技术学院;内蒙古大学生命科学学院)
【作者单位】晋中学院生物科学与技术学院;内蒙古大学生命科学学院
【刊名】晋中学院学报
【关键词】 植物群落;物种丰富度;α多样性;β多样性
【ISSN号】1673-1808
【页码】P56-63
【年份】2019
【期号】第3期
【摘要】在乌金山适合地段沿海拔梯度设置11个样地,采用数量分类,对乌金山植物群落类型、物种组成和物种多样性的垂直分布格局进行调查.结果表明:(1)随着海拔的升高,森林群落依次为油松林、侧柏林、油松侧柏混交林、油松山桃山杏混交林、白皮松林、油松白皮松混交林.(2)11个样地中共调查到乔木13种,灌木16种,草本31种.(3)物种丰富度成明显的单峰分布格局,最大的丰富度出现在中海拔群落中.木本植物的物种丰富度随着海拔高度的增加而下降,草本植物的物种丰富度随着海拔高度的增加先增加后下降.(4)沿海拔梯度森林群落的最大树高、最大胸径、平均树高和平均胸径等群落结构均呈单峰分布格局.(5)α多样性沿海拔梯度变化趋势与物种丰富度相似,但没后者明显.β多样性随海拔的升高而有所降低,在低海拔较高.(6)在调查的海拔范围内,海拔梯度是影响植物群落分布的主要因子,坡度坡向为次要因子,海拔比坡度坡向对群落的结构特征、物种丰富度以及(α、β)多样性的影响更大,而在局部尺度上,人为干扰以及小地形导致的生境异质性对群落物种多样性有着重要影响.
【全文】 文献传递
乌金山植物群落物种多样性的垂直分布格局
摘 要:在乌金山适合地段沿海拔梯度设置11个样地,采用数量分类,对乌金山植物群落类型、物种组成和物种多样性的垂直分布格局进行调查.结果表明:(1)随着海拔的升高,森林群落依次为油松林、侧柏林、油松侧柏混交林、油松山桃山杏混交林、白皮松林、油松白皮松混交林.(2)11个样地中共调查到乔木13种,灌木16种,草本31种.(3)物种丰富度成明显的单峰分布格局,最大的丰富度出现在中海拔群落中.木本植物的物种丰富度随着海拔高度的增加而下降,草本植物的物种丰富度随着海拔高度的增加先增加后下降.(4)沿海拔梯度森林群落的最大树高、最大胸径、平均树高和平均胸径等群落结构均呈单峰分布格局.(5)α多样性沿海拔梯度变化趋势与物种丰富度相似,但没后者明显.β多样性随海拔的升高而有所降低,在低海拔较高.(6)在调查的海拔范围内,海拔梯度是影响植物群落分布的主要因子,坡度坡向为次要因子,海拔比坡度坡向对群落的结构特征、物种丰富度以及(α、β)多样性的影响更大,而在局部尺度上,人为干扰以及小地形导致的生境异质性对群落物种多样性有着重要影响.
关键词:植物群落;物种丰富度;α多样性;β多样性
物种多样性沿环境梯度的变化规律一直是生态学家十分关注的问题.物种多样性不仅能反映群落或生境中物种的丰富度、均匀度和时空变化,表征群落和生态系统的特征及其变化演替的规律,也能反映出不同自然地理条件及人为因素与相关群落的相互关系[1].本文调查乌金山的群落类型、结构及其物种多样性垂直分布格局,运用多样性指数、丰富度指数对乌金山森林群落及林下灌木层和草本层及其灌丛草本群落的物种多样性的分析[2~5].研究物种多样性的梯度格局及控制这些格局的生态因子,是保护生物学研究的基础,其主要与气候、群落生产力和其他因子相关[6~9].调查分析乌金山植物群落的多样性对阐明森林生态系统的形成与维持群落的稳定性与演替规律、种群的生态特征和更新具有极为重要的意义,可为乌金山地区森林景观多样性和植物资源保护提供科学依据.
1 研究地区与研究方法
1.1 研究地区概况
研究区位于乌金山自然保护区,属太行山脉,地理坐标112°46′24″E,37°51′36″N,属暖温带半干旱气候,年均气温为9.8℃.最高海拔为1489.2 m,最低海拔为954.1 m.土壤为砂页岩和砂壤或轻壤.有植物330余种.
1.2 研究方法
采用梯度格局法,海拔每升高50m设置一块样地,共11个.乔木层样方大小为10m×10m;在每个乔木层样方中,选取两个5m×5m的小样方作为灌木层样方,进行灌木调查﹔在每个乔木层样方中梅花取样,设置5个1m×1m草本层样方,对草本层进行调查[10].
1.3 调查内容
(1)样地的基本状况:经纬度、海拔、坡度、坡向等;(2)乔木层中,对胸径>4 cm的所有个体,准确测定其胸径和树高;在灌木层和草本层的调查中,记录每个物种的名称、高度和盖度.
1.3.1 物种多样性测度方法[11]
物种丰富度指数=S;S=样方内所有物种数
Gleason丰富度指数(dm):dm=(S-1)/lnA;A为取样面积
1.3.2 α多样性测度方法
Shannon-Wiener指数(H):H=-∑PilnPi
Pielou均匀度指数(J):J=H/lnS
式中:Pi为样方第i种的重要值;
物种重要值(Pi)的计算:
乔木重要值=(相对盖度+相对频度+相对优势度)/300
灌木植物的重要值=(相对高度+相对盖度)/200
草本植物的重要值=(相对高度+相对盖度)/200
(相对高度就是某一物种高度除以其所在层片的总高度)
1.3.4 β多样性测度方法[5]
β多样性可以定义为沿着环境梯度的变化物种替代的程度.不同群落或某环境梯度上不同点之间的共有种越少,β多样性越大.精确地测定β多样性具有重要的意义.
Jaccard指数CJ=j/(a+b-j)
Cody指数βc=[g(H)+l(H)]/2=(a+b-2j)/2
式中:a、b分别为两群落的物种数,j为两群落的共有物种数,g(H)为沿生境梯度H增加的物种数,l(H)为沿生境梯度H失去的物种数.
2 结果与分析
2.1 乌金山森林群落种类组成分析
由表1可以看出,乌金山森林群落物种丰富,层次分明,成层现象突出,具有明显的垂直结构,分为乔木层、灌木层和草本层.
乔木层有乔木13种,其中油松存在于每个海拔梯度层,分布均匀,数量较多,由表1可以看出油松为乔木层的优势种,对乌金山森林群落的结构和环境的形成有明显的控制作用.灌木层约有16种,其中三亚绣线菊、荆条、黄蔷薇在灌木层中占优势,在一定程度上影响着群落的性质及控制着群落的环境,在中海拔出现较多物种.草本层大约有31种.
表1 乌金山植物群落物种组成

样方 乔木层种类组成 灌木层种类组成 草本层种类组成1(950m)油松(Pinus tabulaeformis)臭椿(Ailanthus altissima)山杏(Armeniaca sibirica)侧柏(Platycladus orientalis)榆树(Ulmus pumila)山桃(Amygdalus davidiana)旱柳(Salix matsudana)杜梨(Pyrus betulifolia)山杨(Populus davidiana)三桠绣线菊荆条(Vitex negundo)枸杞 沙棘(Hippophae rhamnoides)黄蔷薇(Rosa hugonis)小叶鼠李(Rhamnus parvifolia)铁杆蒿(Tripolium vulgare)狗尾草(Setaria viridis)萎陵菜(Potentilla chinensis)针茅(Stipa capillata)猪殃殃(Galium aparine)
表1 乌金山植物群落物种组成(续表)

样方 乔木层种类组成 灌木层种类组成 草本层种类组成2(1000m)油松臭椿侧柏辽东栎(Quercuswutaishanica)山杏旱柳三桠绣线菊(Spiraea salicifolia)荆条 虎榛子(Ostryopsis davidiana)沙棘枸杞黄蔷薇铁杆蒿 苔草(Vittaria flexuosa)萎陵菜长芒草(Stipa bungeana)3(1050m) 油松 五角枫(Acermono)侧柏 臭椿辽东栎杜梨连翘(Forsythia suspensa)多花枸子(Lycium chinense)荆条黄蔷薇小叶鼠李酸枣(Ziziphus jujuba)铁杆蒿 狗尾草山棉花(Thespesia lampas)苔草 山菊(Ser.Oreastra)歪头菜(Vicia unijuga)石竹(Dianthus chinensis)猪殃殃4(1100m)油松臭椿侧柏鹅耳栎(Carpinus turczaninowii)五角枫旱柳三桠绣线菊荆条酸枣虎榛子多花枸子黄蔷薇 小叶鼠李地榆(Sanguisorba officinalis)针茅 棘豆(Oxytropis)山棉花 唐松草(Thalictrum aquilegifolium)苔草 萎陵菜石竹5(1150m) 油松臭椿白皮松五角枫侧柏虎榛子荆条多花枸子黄刺条(Caragana frutex)葱皮忍冬枸杞三桠绣线菊山棉花 针茅 火绒草(Leontopodium leontopodioides)地榆长芒草 棘豆6(1200m) 油松臭椿辽东栎白皮松(Pinus bungeana)侧柏胡枝子多花枸子沙棘 荆条黄刺条六道木(Abelia biflora)葱皮忍冬 三桠绣线菊 河朔荛花(Wikstroemia chamaedaphne)火绒草 糙苏(Phlomis umbrosa)黄背草(Themeda japonica)大丁草(Gerbera anandria)玉竹(Polygonatum odoratum)唐松草黄精(Polygonatum sibiricum)斑叶堇菜(Viola variegata)7(1250m)油松五角枫山杏辽东栎侧柏山桃(Amygdalus davidiana)白皮松荆条 沙棘 胡枝子(Lespedeza bicolor)虎榛子黄刺条葱皮忍冬(Lonicera ferdinandii)六道木蒙古荚蒾(Viburnum mongolicum)小叶鼠李三桠绣线菊黄精 玉竹 小红菊(Dendranthema chanetii)针茅大丁草大蓟(Cirsium japonicum)蒙古堇菜(Violamongolica)唐松草棘豆青杞(Solanum septem lobum)长芒草穿龙薯蓣(Dioscorea nipponica)8(1300m) 油松 鹅耳栎山杏白皮松侧柏山桃三桠绣线菊沙棘虎榛子黄刺条蒙古荚蒾百里香 翻白草针茅歪头菜长芒草棘豆10(1400m) 油松侧柏五角枫白皮松 三桠绣线菊沙棘胡枝子荆条 百里香翻白草(Potentilla discolor)针茅歪头菜铁杆蒿长芒草棘豆糙苏唐松草大丁草小红菊 玉竹百里香(Thymus mongolicus)翻白草大蓟斑叶堇菜9(1350m) 油松五角枫侧柏白皮松鹅耳栎 沙棘虎榛子荆条胡枝子葱皮忍冬11(1450m) 油松 侧柏 山杏 三桠绣线菊沙棘荆条 酸枣枸杞 沙参针茅(Stipa capillata)百里香棘豆糙苏针茅猪殃殃
2.2 物种丰富度与海拔的关系
物种丰富度(species richness)随海拔梯度的变化格局一般有两种:物种丰富度随海拔升高而递减和中海拔地区最丰富.乌金山植物物种丰富度属于在中海拔地区较丰富,中海拔地区降水和温度条件适宜大多数物种的生存,中海拔地区生境较均匀,出现较多种类的草本与灌木,致使植物群落的多样性增加.
从表2,结合图1,2可见,物种丰富度成明显的单峰分布格局,木本植物的丰富度随海拔的升高而呈现递减趋势,而草本植物的丰富度对海拔的升高先增加后下降.物种丰富度在1250m海拔处呈现最高值.中海拔地区降水和温度条件适宜大多数物种的生存,每100m2的样方内一般有17种木本植物以及14种草本植物.
表2 乌金山植物群落物种丰富度随海拔的变化
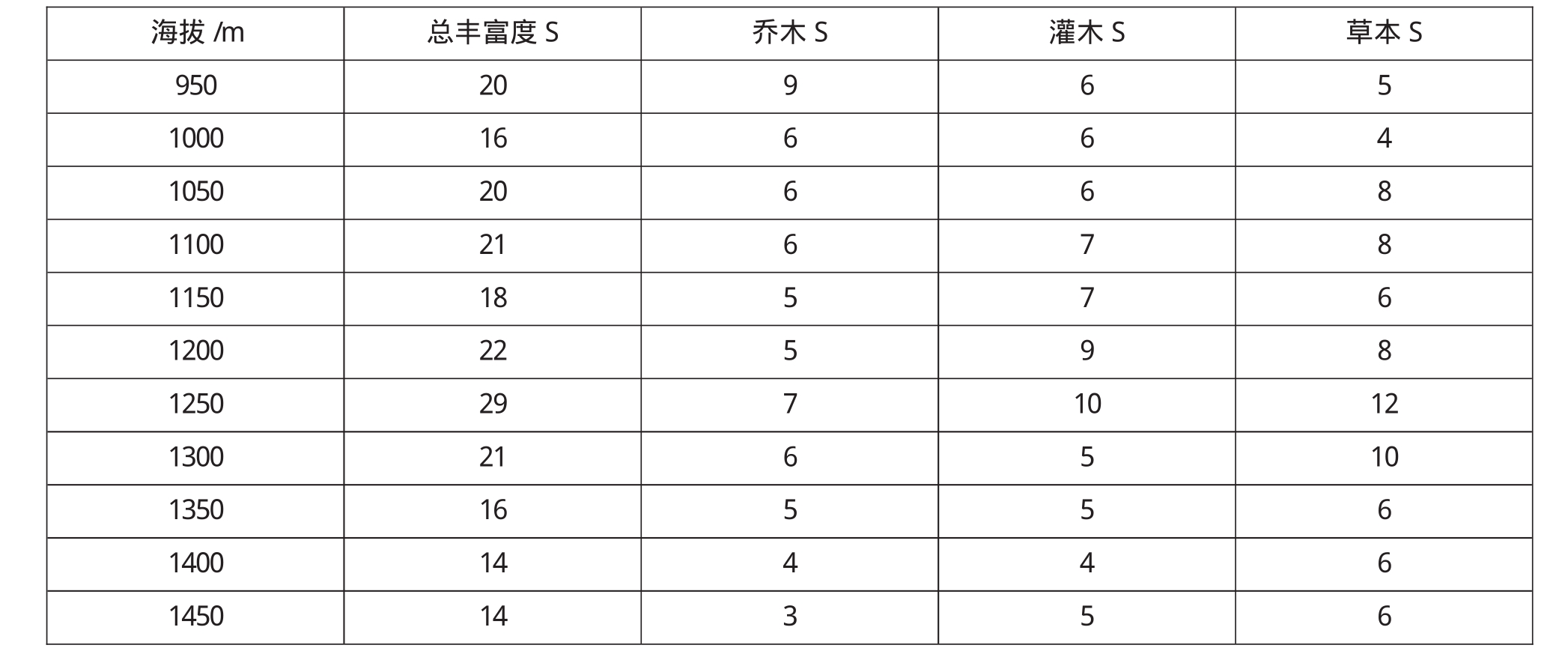
图1 乌金山植物群落物种丰富度随海拔变化
由于不同生活型的植物存在不同的多样性分布格局,本文对乔灌草的丰富度也做了调查,如图2所示,乔木物种数在低海拔地区丰富度较高,随着海拔的升高出现下降趋势,而灌木物种数先增加后减少,因此出现木本植物的物种多样性随着海拔高度的增加而下降.而草本植物随海拔的升高先增加后减少,在中海拔出现较多的草本植物,这与中海拔地区的降水、生境比较均匀,适宜多种物种生存有关.由图2可以看出,乌金山植物群落物种多样性对海拔的敏感性由大到小的次序依次为草本层、灌木层、乔木层.
2.3 群落结构特征
本文群落的结构特征主要通过最大树高、最大胸径、平均树高、平均胸径等4个指标进行分析[8].由表3,图3中可以看出,随着海拔的升高,最大树高(Hmax)、最大胸径(DBHmax)、平均树高和平均胸径等指标总体上呈单峰分布格局.最大树高和最大胸径均在1200m~1300m呈现最高分布.平均胸径与平均树高单峰分布也比较明显.不同生活型的结构特征反应群落的潜在生产力随海拔而变化.
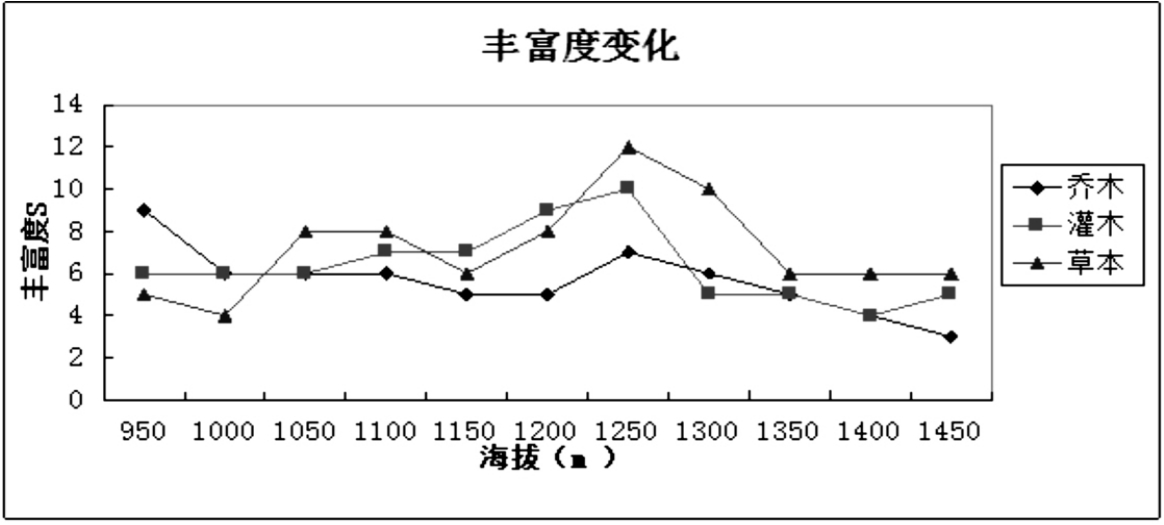
图2 乔灌草本植物物种丰富度随海拔的变化
表3 乌金山植物群落结构特征随海拔的变化格局
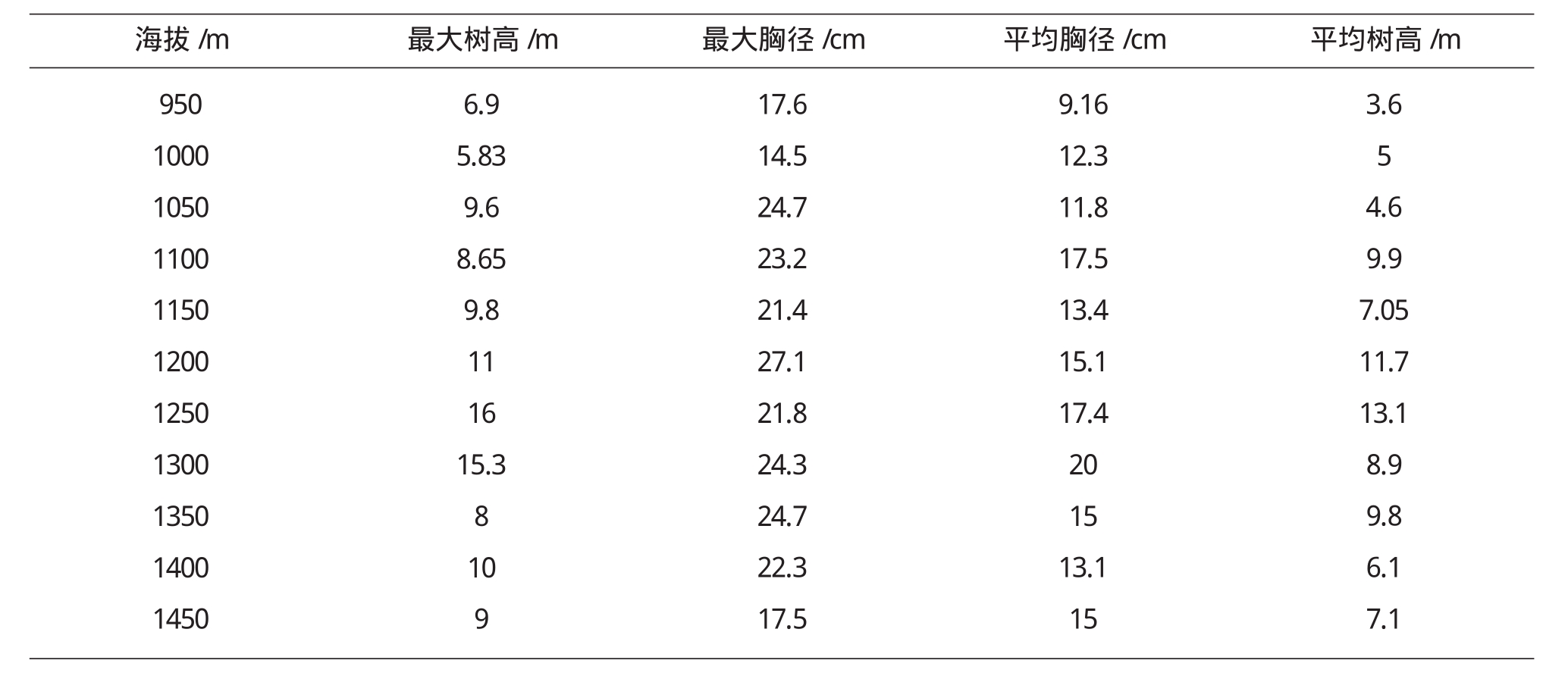
海拔/m最大树高/m最大胸径/cm平均胸径/cm平均树高/m 95010001050110011501200125013001350140014506.95.839.68.659.8111615.3810917.614.524.723.221.427.121.824.324.722.317.59.1612.311.817.513.415.117.4201513.1153.654.69.97.0511.713.18.99.86.17.1
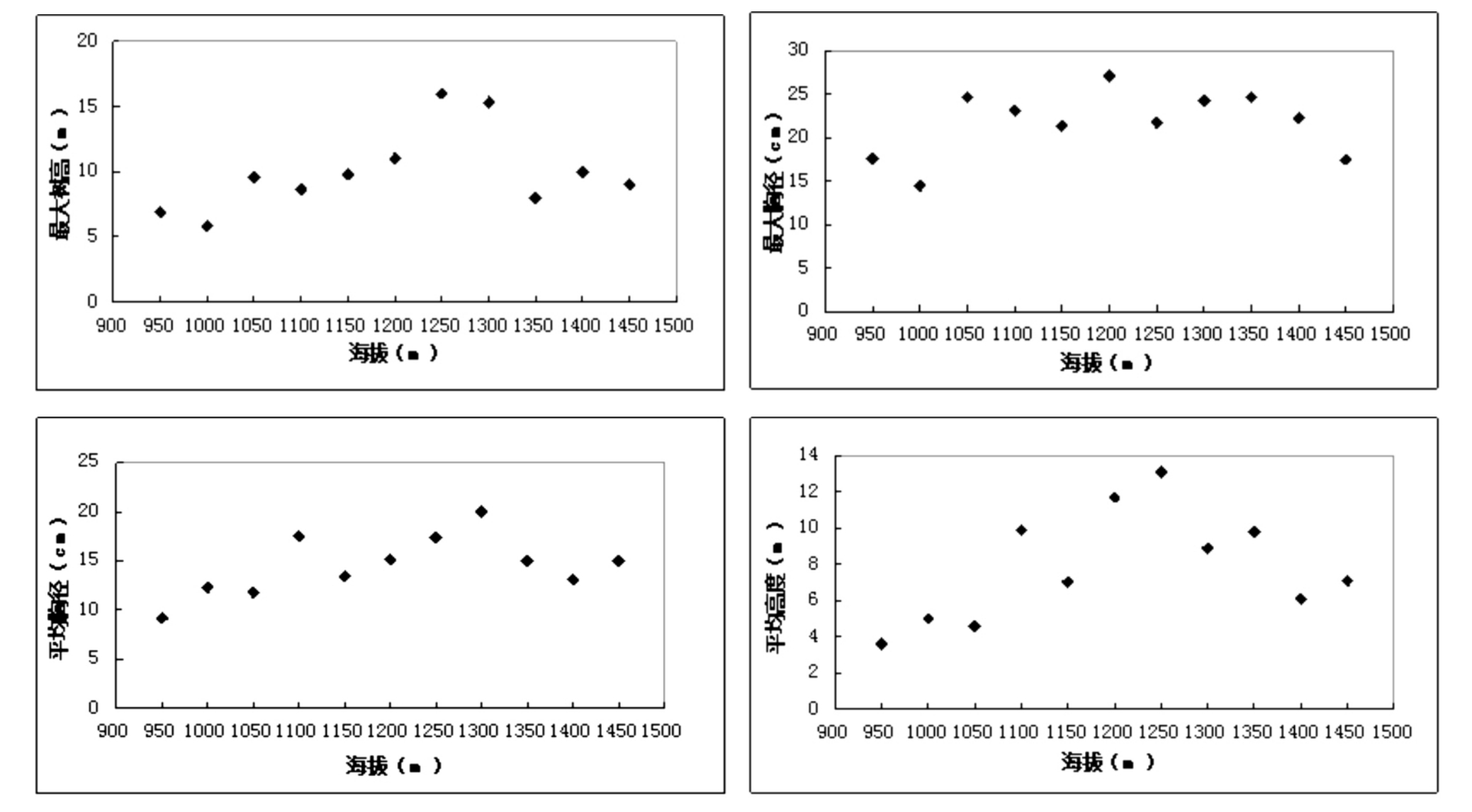
图3 乌金山植物群落结构特征随海拔的变化格局
2.4 α,β多样性随海拔的变化格局
α多样性主要关注局域均质生境下的物种数目,因此被称为生境内的多样性.一般受到小环境以及生物之间的相互作用的影响.本文对α多样性是通过Gleason丰富度指数,Shannon-Wiener指数,Pielou均匀度指数来分析的.
从表4可以看出Shannon-Wiener指数在海拔1250m处达到最大值,结合图1和图4,α多样性随着海拔梯度的变化与物种丰富度相似,随着海拔的升高先增加后降低.这与乌金山的气候条件密切相关,乌金山属于半干旱地区.在低海拔区域,物种多样性受降水的影响,而在高海拔区域受热量的限制,从而形成物种多样性在中海拔出现峰值.
β多样性指沿环境梯度不同生境群落之间物种组成的相异性或物种沿环境梯度的更替速率,即生境间的多样性.一般β多样性随海拔的升高而有所降低,在低海拔处较高,主要原因与干扰及物种之间相互竞争有关.本文是通过Jaccard指数、cody指数来分析β多样性的.Jaccard指数反映群落间物种组成的相似性;Cody指数则反映群落物种组成沿环境梯度的替代速率.
从表5可以看出,Jaccard指数在低海拔地区出现最大值说明群落间物种组成的相似性较大,然后随着海拔的升高而有所降低.结合图5可以看出,在高海拔1350m~1400m出现最大值,此区域属于群落交错区,具有较高的相似性.Cody指数在中海拔区域(1200m~1250m)达到最大值,说明群落物种组成沿环境梯度的替代速率最大.群落相异性表示沿海拔梯度相邻样地间在物种组成上的差异,物种更替则体现物种的替换速率或数量.
表4 乌金山沿海拔梯度Shannon-wiener指数和均匀度指数的变化

海拔/m shannon-wiener指数H均匀度指数J 95010001050110010501200125013001350140014502.32.82.53.12.62.93.22.62.721.80.911.10.730.780.690.820.570.640.530.60.55
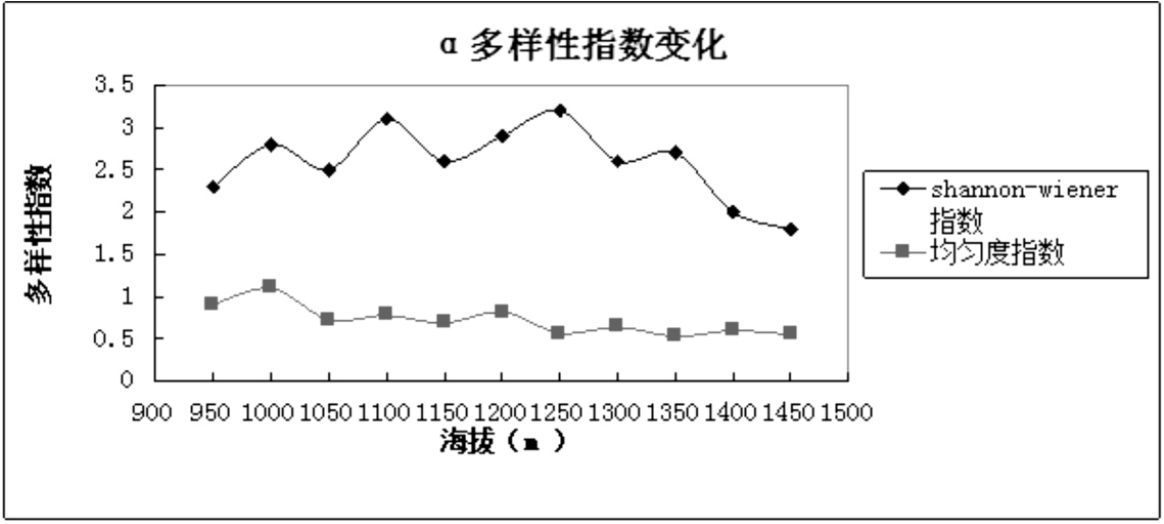
图4 乌金山沿海拔梯度Shannon-w iener指数和均匀度指数的变化
表5 乌金山沿海拔梯度相邻群落间Jaccard指数和Cody指数
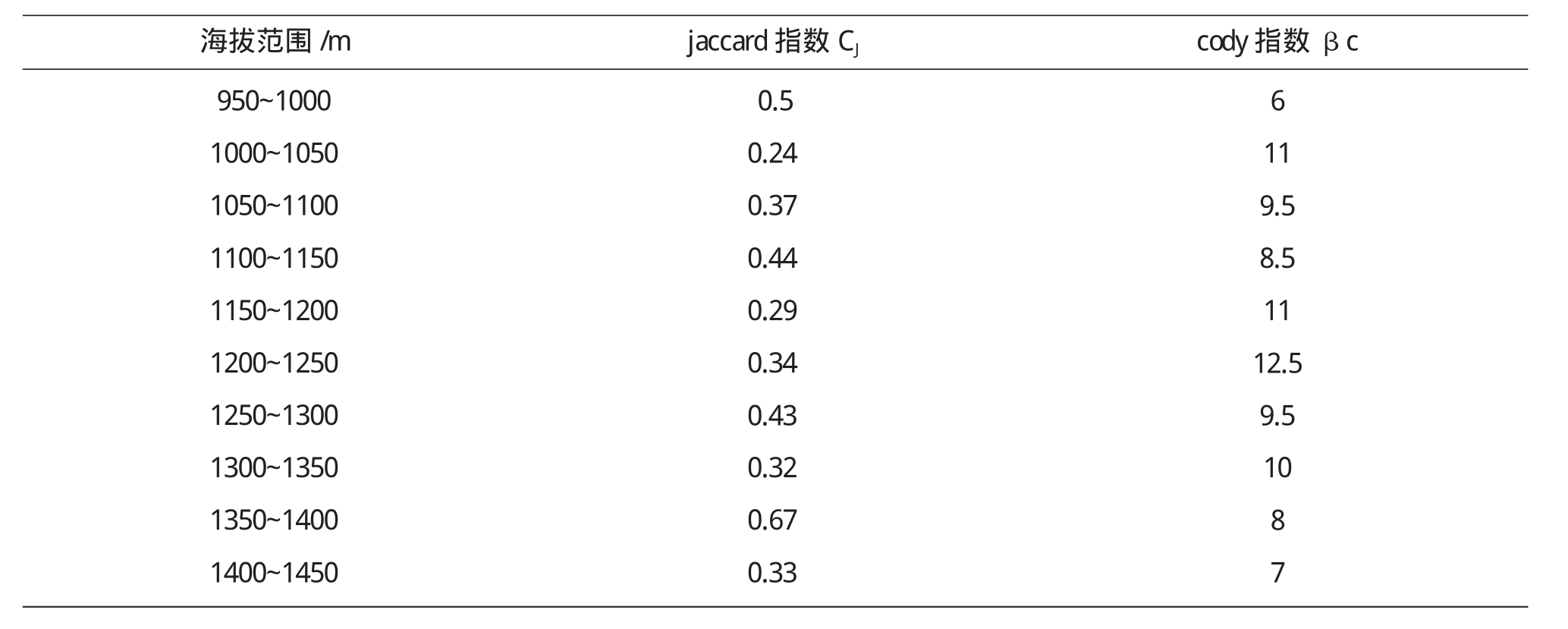
海拔范围/m jaccard指数CJ cody指数βc 950~10001000~10501050~11001100~11501150~12001200~12501250~13001300~13501350~14001400~14500.50.240.370.440.290.340.430.320.670.336119.58.51112.59.51087
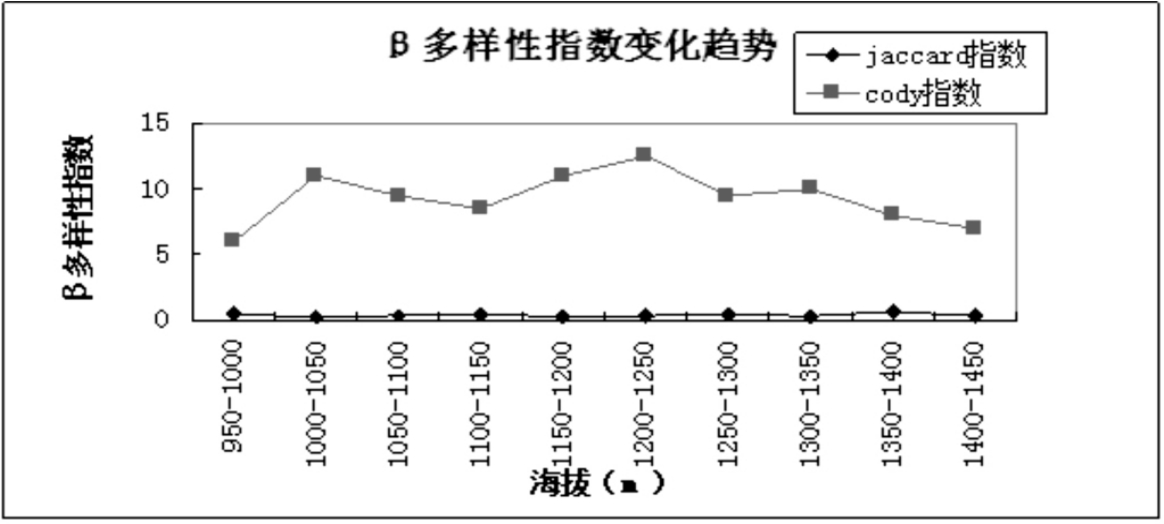
图5 乌金山沿海拔梯度相邻群落间Jaccard指数和Cody指数
3 讨论
由调查可以看出乌金山植物群落物种种类丰富,垂直结构明显,共调查到乔木层有13种,灌木层有16种,草本层大约有31种.随着海拔的升高,森林群落依次为油松林、侧柏林、油松侧柏混交林、油松山桃山杏混交林、白皮松林、油松白皮松混交林.水分和能量对植物物种多样性有重要的影响.在中海拔群落(1200m~1250m)物种丰富度出现最大的值,在中海拔群落,水分、能量比较充足,适宜大多数物种生存,出现较多种类的草本与灌木,致使植物群落的多样性增加.另外也与人为因素有关.在低海拔处人为活动频繁,有损生物多样性,在低海拔区域的物种多样性受降水的影响,而在高海拔区域受热量的限制,从而形成物种多样性在中海拔出现峰值.木本植物的物种丰富度随着海拔高度的增加而下降,草本植物的物种丰富度随着海拔高度的增加先增加后下降.乌金山植物群落物种多样性对海拔的敏感性由大到小的次序依次为草本层、灌木层、乔木层.这与张峰、张金屯、上官铁梁对历山自然保护区猪尾沟森林群落植物多样性研究所得结论有所差异,主要原因是乌金山与历山所处地理位置不同、环境因子不同、生态气候不同,从而使不同生活型的物种多样性出现了差异.
α多样性一般受到小环境和生物之间的相互作用的影响.α多样性随着海拔梯度的变化与物种丰富度相似,随着海拔的升高先增加后降低.这与乌金山的气候条件密切相关,乌金山属于半干旱地区.在低海拔区域,物种多样性受降水的影响,而在高海拔区域受热量的限制,从而形成物种多样性在中海拔出现峰值.α多样性主要与水热条件有关,符合唐志尧、方精云的植物物种多样性的垂直分布格局研究的α多样性一般格局.影响β多样性的主要因子有土壤、地质以及干扰等,β多样性随海拔的升高而有所降低,在低海拔较高,主要原因与干扰及物种之间相互竞争有关,低海拔生境异质性较大,在土壤和地形变化频繁的地段,群落含更多植物种.因为海拔不同出现不同的群落,在群落交错的地段,物种多样性较大.
参考文献
[1]王国宏.祁连山北坡中段植物群落多样性的垂直分布格局[J].生物多性,2002(10):7~14.
[2]高贤明,陈灵芝.北京山区辽东栎群落物种多样性的研究[J].植物生态学报,1998(22):23~32.
[3]唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20~28.
[4]曾从盛.福建典型区生态环境研究[M].北京:中国环境科学出版社,2006.
[5]朱彪,陈安平,刘增力,等.广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局[J].生物多样性,2004,12(1):44~52.
[6]徐东成,冯建孟,王襄平,等.云南高黎贡山北段植物物种多样性的垂直分布格局[J].生态学杂志,2008,27(3):323~327.
[7]赵淑清,方精云,朴世龙,等.大兴安岭呼中地区白卡鲁山植物群落结构及其多样性研究[J].生物多样性,2004,12(1):182~189.
[8]冯建孟,王襄平,徐成东,等.玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局[J].山地学报,2006,24(1):110~116.
[9]唐志尧,柯金虎.秦岭牛背梁植物物种多样性垂直分布格局[J].生物多样性,2004,12(1):108~114.
[10]方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533~548.
[11]李智琦,欧阳志云,曾慧卿.基于物种的大尺度生物多样性热点研究方法[J].生态学报,2010,30(6):1586~1593.
Vertical Distribution Pattern of Plant Community Species Diversity in M t.W ujin
Abstract:Eleven sampling plots were established along an altitudinal gradient in Mt.Wujin,and the vertical distribution pattern of plant community,species composition,and species diversity were studied by using quantitative classification.The results show that(1)with the increase of altitude,the forest communitiesweremixed forest of Chinese pine forest,side of Berlin,Chinese pine and arborvitaemixed,pine Prunus davidiana wild apricot,White pine,Chinese pine and Pinus bungeanamixed;(2)A total of 13 kinds of trees,15 species of shrubs,and 31 kinds of herbswere recorded;(3)Unimodal distribution pattern of species richness is significant,and the biggest richness appears in themedium altitude.Woody plant species richness declined simultaneously with the increase of altitude,herbaceous plants of the species richness increased with the increase of altitude first and then declined;(4)With the increase of altitude,the maximum height,maximum DBH,average height and average DBH of trees all showed an increasing pattern,and then a decreasing pattern with the increase of elevation;(5)The change trends ofαdiversity along an altitudinal gradient are similar to species richness,but not the latter obviously.βdiversity have a rising trend in low altitude;(6)Elevation within the scope of this study,elevation gradient are the main factors that affect the distribution of plant communities,slope to the secondary factors,on a local scale,human disturbance and habitat heterogeneity on small terrain have an important impacton species diversity.
Keywords:plant community;species richness;αdiversity;βdiversity
中图分类号:Q 948
文献标志码:A
文章编号:1673-1808(2014)03-0056-08
[收稿日期]2013-03-24
曲 波(1984-),男,山西平陆人,晋中学院生物科学与技术学院,助教,硕士,研究方向:环境生态学、植被生态学;
牛霞霞(1990-),女,山西高平人,内蒙古大学生命科学学院,硕士,研究方向:植被生态学.
book=443,ebook=443