
红脂大小蠹种群空间格局地统计学分析及抽样技术
【类型】期刊
【作者】潘杰,王涛,宗世祥,温俊宝,骆有庆(北京林业大学省部共建森林培育与保护教育部重点实验室;北京市门头沟林业站)
【作者单位】北京林业大学省部共建森林培育与保护教育部重点实验室;北京市门头沟林业站
【刊名】生态学报
【关键词】 红脂大小蠹;混交林;空间格局;地统计学;空间抽样
【资助项】北京市教育委员会共建项目(JD100220888);中央高校基本科研业务费专项资金资助
【ISSN号】1000-0933
【页码】P195-202
【年份】2019
【期号】第1期
【期刊卷】1;|7;|8;|4;|5;|2
【摘要】红脂大小蠹是我国一重要林业外来入侵害虫,主要危害油松、白皮松和华山松,目前已扩散到山西、陕西、河北、河南、北京等地。为了研究其种群空间格局,用地统计学方法分析了受害程度不同的油松纯林和混交林林分内红脂大小蠹种群空间分布特性。结果表明:两种受害程度不同的油松纯林内红脂大小蠹的危害具有显著差异,轻度受害林分内有虫株率为4.9%,而重度受害林分则达到23.6%。全方向的变异函数曲线图分析得知:轻度受害纯林、重度受害纯林、混交林林分内红脂大小蠹种群的空间依赖范围分别为49.400、47.400、73.820m,而局部空间连续性强度分别为0.824、0.582、0.762。轻度受害纯林与重度受害纯林林分内红脂大小蠹种群均表现为较强的空间聚集性,而混交林受害林分内其种群则表现为随机分布。受害程度不同的纯林林分内红脂大小蠹种群空间格局具有一定的差异,而且纯林与混交林林分内其种群空间格局也有较大的差别。本文根据红脂大小蠹种群空间分布特性,对其种群空间抽样技术与传统抽样技术做了相应的比较与探讨。
【全文】 文献传递
红脂大小蠹种群空间格局地统计学分析及抽样技术
摘要:红脂大小蠹是我国一重要林业外来入侵害虫,主要危害油松、白皮松和华山松,目前已扩散到山西、陕西、河北、河南、北京等地。为了研究其种群空间格局,用地统计学方法分析了受害程度不同的油松纯林和混交林林分内红脂大小蠹种群空间分布特性。结果表明:两种受害程度不同的油松纯林内红脂大小蠹的危害具有显著差异,轻度受害林分内有虫株率为4.9%,而重度受害林分则达到23.6%。全方向的变异函数曲线图分析得知:轻度受害纯林、重度受害纯林、混交林林分内红脂大小蠹种群的空间依赖范围分别为49.400、47.400、73.820m,而局部空间连续性强度分别为0.824、0.582、0.762。轻度受害纯林与重度受害纯林林分内红脂大小蠹种群均表现为较强的空间聚集性,而混交林受害林分内其种群则表现为随机分布。受害程度不同的纯林林分内红脂大小蠹种群空间格局具有一定的差异,而且纯林与混交林林分内其种群空间格局也有较大的差别。本文根据红脂大小蠹种群空间分布特性,对其种群空间抽样技术与传统抽样技术做了相应的比较与探讨。
关键词:红脂大小蠹;混交林;空间格局;地统计学;空间抽样
Abstract:Red turpentine beetle(RTB),Dendroctonus valens LeConte was a destructive forest invasive species in China,it mainly attacks Pinus.tabuliformis and P.bungeana.So far it has spread rapidly to the provinces of Shanxi,Hebei,Henan and Beijing since its first outbreak in Shanxi province in 1998,and caused extensive tree mortality.In order to further study the spatial distribution of its population,the spatial characteristics of D.valens population were analyzed by using geostatistical methods in the pure P.tabuliformis forest and mixedwood stands which at different damage levels.The result showed that the damage level caused by D.valens differed significantly in the two woodlands.Only 4.9%of trees in lightly-infested woodland were infested compared with 23.6%infested individuals in heavily-infested woodland.Analysis of the isotropic variogram indicated that the distance of spatial dependence of D.valens was 49.400m in the lightly-infested levels of pure P.tabuliformis forest,47.400m in the heavily-infested one and 73.820m in the mixedwood stands,while the intensity of local spatial continuity was 0.824,0.582,0.762 respectively in the above stands.Intense spatial aggregation distribution pattern of D.valens were observed in the lightly-infested level and heavily-infested level of pure P.tabuliformis forest,while random distribution was observed in the mixedwood forest.It concluded that the difference of spatial distribution of D.valens not only existed in different damage levels of pure forest,but also in different stands types. According to the spatial distribution of its population,the spatial sampling technique of D.valens was also studied,and then compared with traditional sampling technique.
Key Words:Dendroctonus valens;mixedwood stands;spatial distribution pattern;geostatistics;spatial sampling
红脂大小蠹(Dendroctonus valens LeConte)又名强大小蠹,属鞘翅目(Coleoptera)小蠹科(Scolytidae),在我国为一新记录外来入侵种。国外分布于美国、加拿大、洪都拉斯等国,危害各种松树,有时也危害云杉和冷杉[1-3]。在中国,1998年在山西阳城、沁水首次发现该虫,目前已知主要寄主为油松、白皮松和华山松。截止2004年底,红脂大小蠹已扩散到山西、陕西、河北、河南4省,发生面积超过5×105hm2,枯死松树达600多万株[1,4],2005年又扩散至北京市门头沟区[5]。
空间格局是昆虫种群重要属性之一,由物种生物学特性与环境条件所决定。研究空间格局有助于了解昆虫生态特性,对揭示种群空间结构、提高抽样技术、种群消长趋势和扩散范围的预测预报、制订害虫防治策略等都具有重要意义[6-7]。苗振旺、张历燕、李建康等先后都对红脂大小蠹种群空间格局及传统抽样技术进行了研究[8-10],但传统的空间格局分析及抽样技术忽略了研究对象的空间位置,得到的结果很大程度上依赖于取样单位的大小,无法反映其局部变化特征,且研究都是基于油松纯林进行的,并未探讨混交林内的情况;而空间抽样技术则可以避免以上问题,不仅能考虑抽取多少样本,而且还能兼顾在多大范围内抽取,以及在空间的什么地方抽取等问题,从而制定更为科学的抽样方案。本文应用地统计学方法[11-16]对油松纯林与混交林林分内红脂大小蠹种群空间格局及空间抽样技术进行较深入研究,并对空间抽样技术与传统抽样技术做了相应比较与探讨,以期为种群预测预报以及采取适宜的防治方法等提供理论指导。
1 试验地概况
试验地设在山西省太岳山国有林管理局灵空山林场,位于太岳林区中部,沁源县西南部,地理位置为东经112°02'—112°09',北纬36°31'—36°43',平均海拔l500m,属暖温带大陆性季风气候,年均气温8℃,年降雨量650mm,年平均相对湿度65%。植被类型包括:未经改造的天然次生林,以辽东栎、油松纯林为主,还有少量杨桦林;近几年新造的油松、落叶松幼林;以虎榛子、胡枝子、沙棘、山桃、黄刺梅为主的灌木林[17]。
2009年7—8月在灵空山林场选取3块林地作为试验地,包括2块受害程度不同的纯林和1块混交林。
(1)轻度受害油松纯林 土壤腐殖质丰富,郁闭度0.9,平均胸径18.9cm,树高9.6m。油松受害率4.9%。
(2)重度受害油松纯林 土壤较贫瘠,郁闭度0.7,油松平均胸径19.3cm,树高9.8m。油松受害率23.6%。
(3)混交林 树种以油松、辽东栗、胡枝子为主,土壤较疏松,土质较好,郁闭度0.9,油松平均胸径为17.4cm,树高8.7m。油松受害率7.5%。
2 研究方法
2.1 调查方法
采取逐株取样调查的方法,统计每株油松上当年侵入孔数量并以此作为密度指标,即每株受害油松上的成虫虫口数(头/株),同时记录油松株行距,将每株油松的相对空间位置绘制于坐标纸上,为半方差函数分析及空间抽样技术提供参考与支持。
2.2 分析方法
2.2.1 空间格局分析
(1)半方差函数
半方差函数是指区域化变量Z(xi)和Z(xi+h)的增量平方的数学期望,即区域化变量增量的方差。半方差函数既是距离h的函数,又是方向α的函数。其公式如下所示:
式中,γ(h)为半方差函数值,半方差函数曲线图是半方差函数γ(h)对距离h的坐标图形。N(h)是被h分隔的数据对的数量,Z(xi)和Z(xi+h)分别是在点xi和xi+h处样本的测量值,h是两分隔样点的距离。
应用半方差函数分析种群空间格局,实质上是分析表征种群数量的变量空间变异特征和程度。计算区域化变量不同方向、不同距离的半方差函数值并应用一定的模型进行模拟,可以将这些值及其模拟值制成如图1所示的曲线图。
变异曲线图中有3个最重要的参数:块金常数,基台值和变程。块金常数是指变异函数曲线被延伸到间隔为零时的截距,它反映了区域化变量内部随机性的可能程度。基台值是指达到平衡时的变异函数值,它反映了变量变化幅度。变程是当变异函数值达到平衡时的间隔距离,它反映了区域化变量的影响范围。
(2)变异函数理论模型的拟合
拟合实际变异曲线图的理论模型大致有球状模型、指数模型、高斯模型、线性有基台值模型和线性无基台值模型等,选择何种理论模型进行拟合,需要根据判断最优拟合模型的方法,即先考虑决定系数R2大小,然后考虑残差RSS大小,最后再考虑变程和块金值大小来判断不同理论模型在拟合实际变异曲线图时的优劣程度[18]。
2.2.2 空间抽样技术
经典的抽样方法包括简单随机抽样、分层随机抽样、集团随机抽样、系统随机抽样和系统非随机抽样等[19]。如何选择合适的抽样方法取决于各种抽样方法的比较与昆虫的生物学特性等因素。研究表明红脂大小蠹危害是从中心开始扩散至周围,故采取分层随机抽样。其基本原理为:区域D被分为不重叠的层,每层选择简单随机样本。需要考虑层的大小、层的形状和每层中抽样数的多少,一般层的划分可以根据实际情况而定,如考虑对从扩散中心开始的扩散后种群密度的估计,显然可以假定离扩散中心越远密度越低,因此可以选择同心圆面积增大形式的层[19]。具体的抽样步骤如下[20]:
①根据昆虫种群的空间依赖性距离划定抽样范围,一般抽样范围略大于变程。
②确定适当的抽样数,一般情况,空间相关的强弱决定了抽样数的多少,空间变化连续性越强的地方抽样数越多。抽样数的大小也有经验上的考虑,因为抽样太多会带来计算上的麻烦,也未必对空间估计有帮助。
③在划定的范围内采用等概重复方式抽取样本。
④计算样本的半方差函数并与总体的半方差函数比较。
⑤提供达到精度要求的抽样方案(精度要求视具体情况而定);否则回到步骤③。
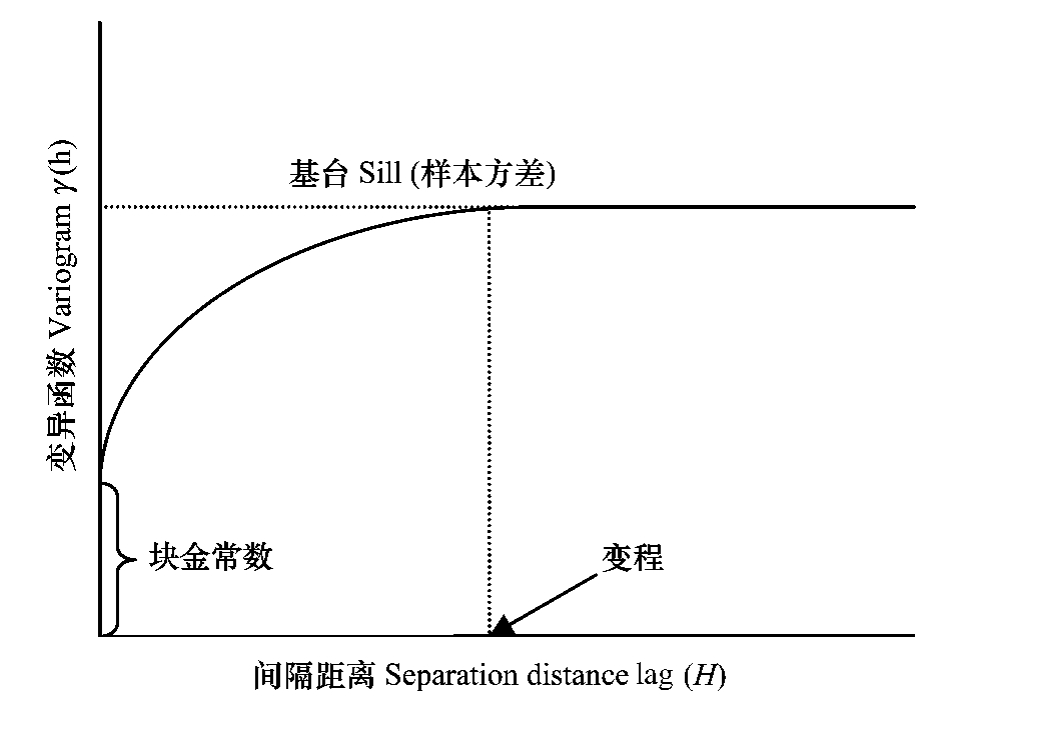
图1 典型的变异曲线
Fig.1 A typical variogram
3 结果与分析
3.1 红脂大小蠹种群空间格局
3.1.1 不同受害程度纯林林分内种群数量分布状况
由图2和图3可以看出两种受害程度不同的油松纯林内,红脂大小蠹的危害具有较大差异,轻度受害林分内有虫株率为4.9%,而重度受害林分则达到23.6%。
3.1.2 红脂大小蠹种群数量的变异函数与空间格局
使用GS+软件,对红脂大小蠹种群空间格局进行地统计学分析。不同受害程度的纯林与混交林内红脂大小蠹种群数量变异函数理论模型各参数及空间格局见表1。
(1)轻度受害纯林林分内红脂大小蠹种群数量空间格局分析
由表1、图4均表明轻度受害油松纯林中的红脂大小蠹种群具有空间依赖性,其空间格局为聚集分布。空间依赖范围为49.400m,即在此距离范围之内,抽样空间上任意两样点种群数量间都存在着一定的相关关系。半方差函数中块金常数占基台值的比例——空间不连续性强度为17.6%,这表明在同一样方内,由于空间依赖性而减少的变异占总体方差的82.4%。块金常数可能是由两方面原因引起的:①试验地内一部分油松被割脂,有可能使红脂大小蠹分布数量相对减少;②试验地内同时进行红脂大小蠹引诱试验,也有可能影响红脂大小蠹的分布。
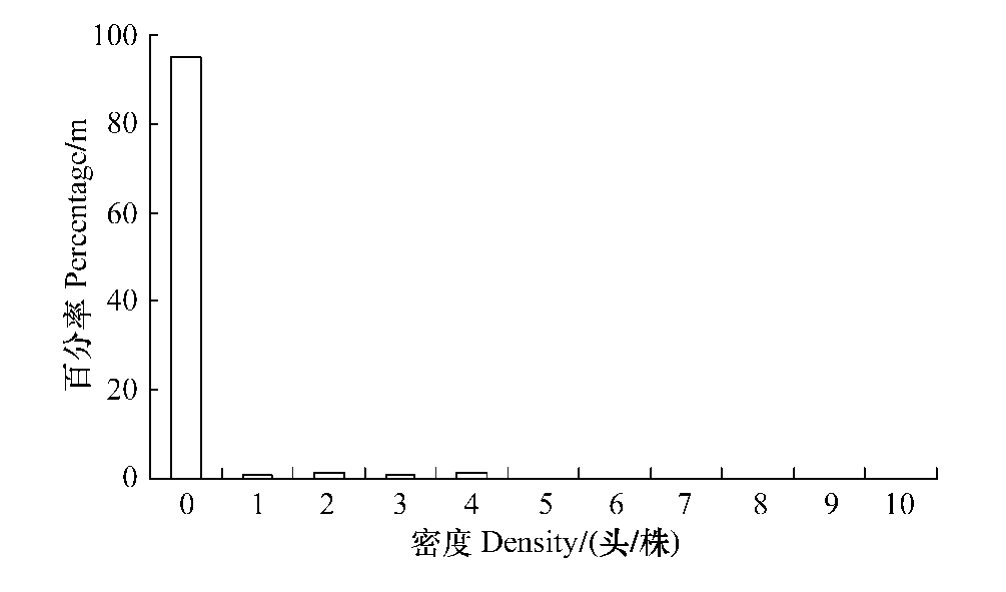
图2 轻度受害林分虫口密度分布
Fig.2 Histogram of density for D.valens in lightly infested stand
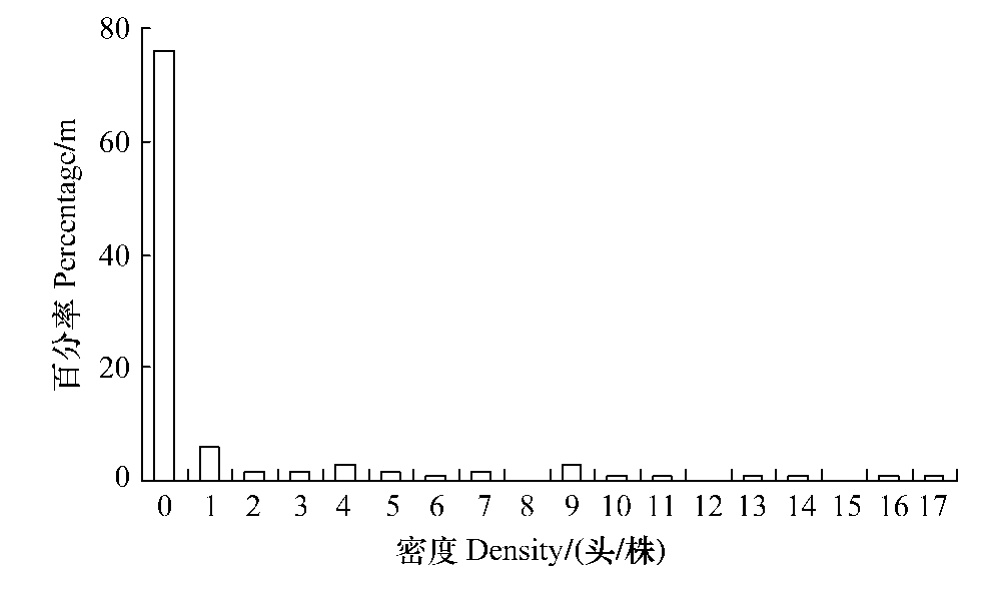
图3 重度受害林分虫口密度分布
Fig.3 Histogram of density for D.valens in heavily infested stand
表1 红脂大小蠹种群数量的变异函数理论模型拟合参数及空间分布型
Table 1 Parameters of variogram models and spatial distribution of the population number for D.valens
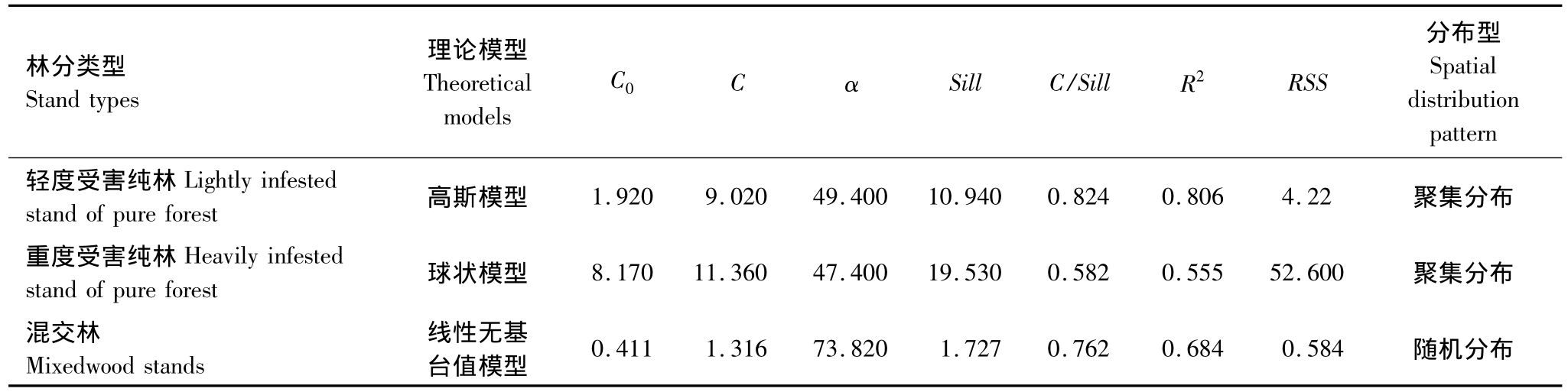
* C0为块金常数,C为拱高,α为变程即空间依赖范围,Sill为基台值,C/Sill为空间变异,R2为决定系数,RSS为残差平方和
林分类型Stand types理论模型Theoretical models C0C α Sill C/Sill R2RSS分布型Spatial distribution pattern轻度受害纯林Lightly infested stand of pure forest 高斯模型 1.920 9.020 49.400 10.940 0.824 0.806 4.22聚集分布重度受害纯林Heavily infested stand of pure forest 球状模型 8.170 11.360 47.400 19.530 0.582 0.555 52.600 聚集分布混交林Mixedwood stands随机分布线性无基台值模型 0.411 1.316 73.820 1.727 0.762 0.684 0.584
由红脂大小蠹种群数量的等值线与矢量叠加图(图4)可以看出:样地内红脂大小蠹分布呈聚集状态。整个区域分布较零星,但是图中东南方向红脂大小蠹分布较集中,密度较大。原因是为了减轻红脂大小蠹对油松的危害,在样地的边缘悬挂了大量的诱捕器。这就导致红脂大小蠹向林地边缘聚集,使虫口密度急剧升高。
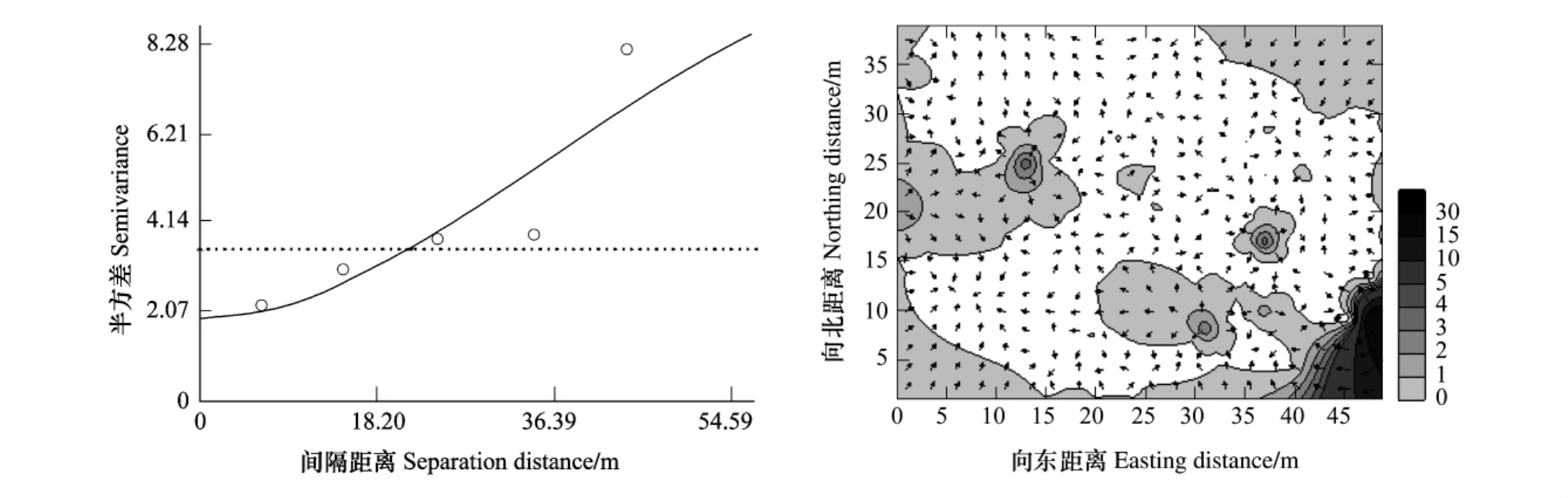
图4 轻度受害纯林林分中红脂大小蠹种群数量的变异函数图(a)和等值线与矢量叠加图(b)
Fig.4 Variogram(a),overlay map of isoline and vector(b)of the population number for D.valens in lightly infested stand of pure P.tabuliformis forest
(2)重度受害纯林林分内红脂大小蠹种群数量空间格局分析
由表1、图5可以看出重度受害油松纯林林分中,红脂大小蠹种群也呈现出较明显的空间聚集状态,其空间依赖范围为47.400m,即在此距离范围内任何两样点种群数量都存在一定的相关关系,说明红脂大小蠹种群在空间上的分布具有连续性,其强度随距离增加而减小。变异函数图中拱高C所占比例为58.2%,说明总空间变异中有58.2%是由空间自相关现象产生。块金常数较大,原因:①重度危害纯林中有一部分油松枯死,某些油松的枯死造成数据缺失,红脂大小蠹虫口数据只能以0计数,这样就与邻近的样方产生较大的差异;②由于人为调查误差造成某些侵入孔未被统计在内;③试验地内同时进行红脂大小蠹引诱试验,也有可能影响红脂大小蠹的分布。
由红脂大小蠹种群数量的等值线与矢量叠加图(图5)可以看出,其密度的聚集分布特点,基本上呈斑块状分布,整个区域内有若干聚集点,而每个聚集点则分别由聚集点中心区域向四周逐渐减小。图中也可以看出样地南边红脂大小蠹虫口密度较大,原因是样地的边缘悬挂了很多诱捕器,从而导致红脂大小蠹大量聚集。
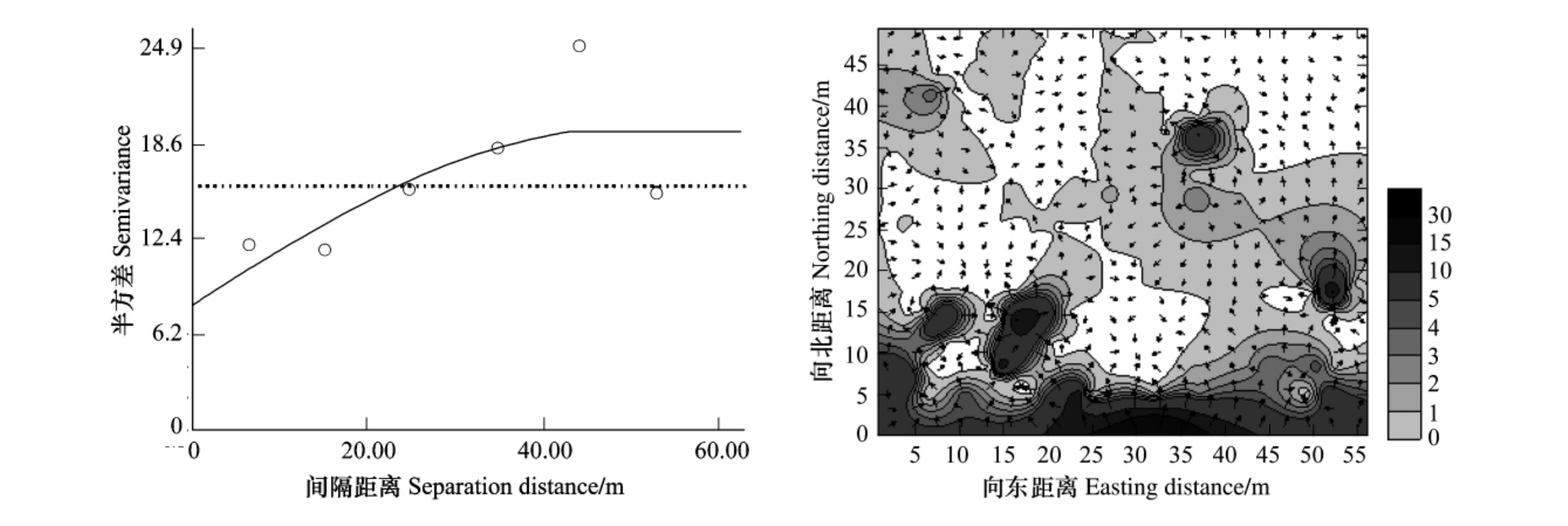
图5 重度受害纯林林分中红脂大小蠹种群数量的变异函数图(a)和等值线与矢量叠加图(b)
Fig.5 Variogram(a),overlay map of isoline and vector(b)of the population number for D.valens in heavily infested stand of pure P.tabuliformis forest
(3)混交林林分内红脂大小蠹种群数量空间格局分析
由表1、图6可以看出,油松混交林林分中红脂大小蠹种群呈现较弱的聚集性,基本上接近于随机分布。由其变异函数曲线图可知其空间依赖范围为73.820m,即在此距离范围之内,抽样空间上任意两样点种群数量间都存在着一定的相关关系。空间变异值为0.762,即在抽样空间种群的随机性较强,这说明在抽样尺度空间中被研究对象的变化较小即种群相互独立性强,空间分布格局是随机分布。块金常数的产生可能由以下因素所引起:混交林内经常有人放牧,人畜活动可能影响红脂大小蠹生活习性。
由红脂大小蠹种群数量的等值线与矢量叠加图(图6)可以看出,混交林内红脂大小蠹基本呈随机化分布。其原因可能是混交林内其他树种阻碍或减弱了红脂大小蠹与油松萜烯类挥发物的信息交流,以至于红脂大小蠹不能明确的寻找寄主,呈现出随机化分布状态。
3.2 空间抽样技术
采取分层随机抽样对几种不同受害类型样地进行抽样估计,各种情况限于篇幅仅以表2作以说明。
①轻度受害纯林林分内红脂大小蠹种群的空间依赖范围为49.400m,可将抽样范围划为30m,即以虫源树(枯死树或濒死树)为中心,半径分别为10、20、30m的范围进行取样。经多次抽样比较,最终确定抽样数为50株时,所抽样本的半方差函数与总体半方差函数的平均误差为10.8%,平均精度为89.2%。
②重度受害纯林林分内红脂大小蠹种群的空间依赖范围为47.400m,可将抽样范围划为30m,即以虫源树(枯死树或濒死树)为中心,半径分别为10、20、30m的范围进行取样。经多次抽样比较,最终确定抽样数为50株时,所抽样本的半方差函数与总体半方差函数的平均误差为6.4%,平均精度为93.6%。
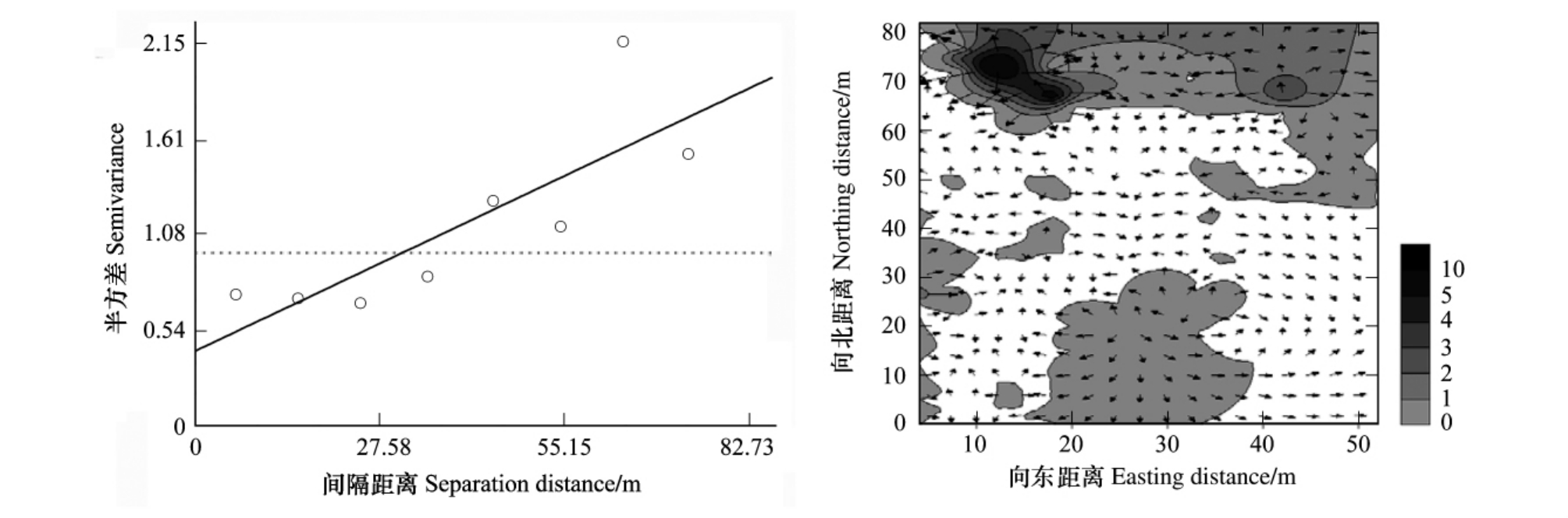
图6 混交林林分中红脂大小蠹种群数量的变异函数图(a)和等值线与矢量叠加图(b)
Fig.6 Variogram(a),overlay map of isoline and vector(b)of the population number for D.valens in mixedwood stands
③混交林林分内红脂大小蠹种群的空间依赖范围为73.820m,可将抽样范围划为40m,即以虫源树(枯死树或濒死树)为中心,半径分别为10、20、30、40m的范围进行取样。经多次抽样比较,最终确定抽样数为50株时,所抽样本的半方差函数与总体半方差函数的平均误差为15.8%,平均精度为84.2%。
表2 红脂大小蠹在不同受害林分中理论抽样数值表
Table 2 The theory sampling number of D.valens in the different degree of affected stands
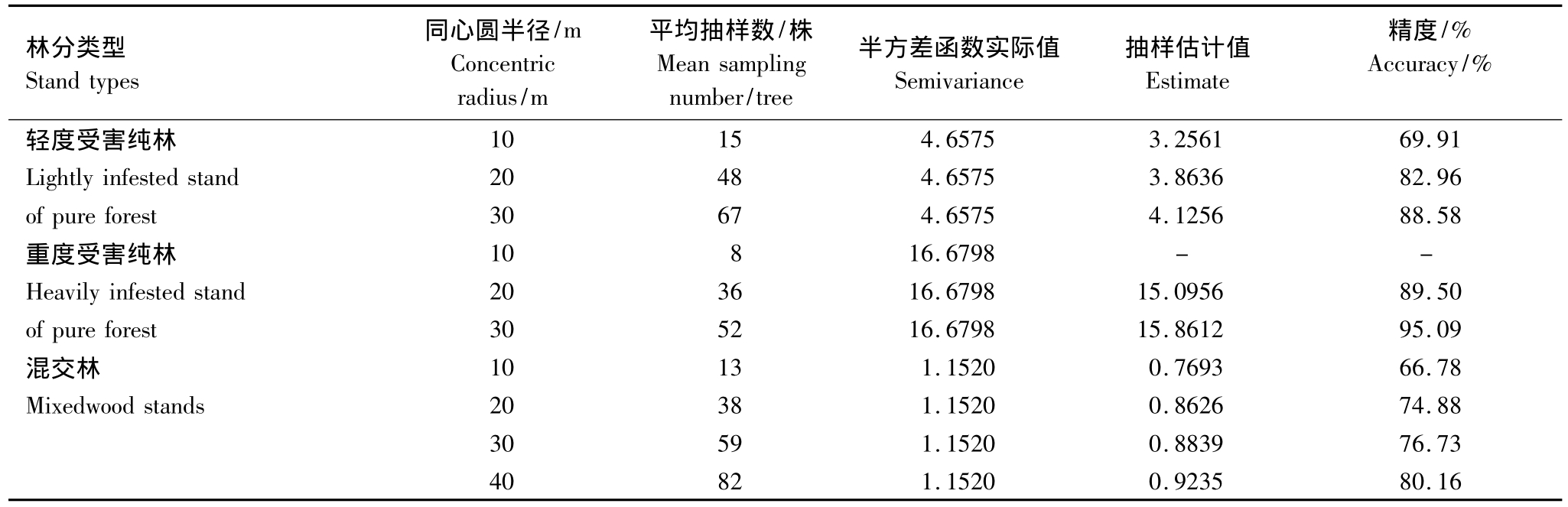
林分类型Stand types同心圆半径/m Concentric radius/m平均抽样数/株Mean sampling number/tree半方差函数实际值Semivariance抽样估计值Estimate精度/% Accuracy/% 10 15 4.6575 3.2561 69.91 Lightly infested stand 20 48 4.6575 3.8636 82.96 of pure forest 30 67 4.6575 4.1256 88.58重度受害纯林 10 8 16.6798 - -Heavily infested stand 20 36 16.6798 15.0956 89.50 of pure forest 30 52 16.6798 15.8612 95.09混交林 10 13 1.1520 0.7693 66.78 Mixedwood stands 20 38 1.1520 0.8626 74.88 30 59 1.1520 0.8839 76.73轻度受害纯林40 82 1.1520 0.9235 80.16
又依据传统空间格局分析方法[21-24]对其种群传统抽样技术进行了研究,结果见下表(表3),以此来对照比较传统抽样技术与空间抽样技术的差异。
表3 红脂大小蠹在不同虫口密度下理论抽样数表
Table 3 The theory sampling number with different D.valens adults density
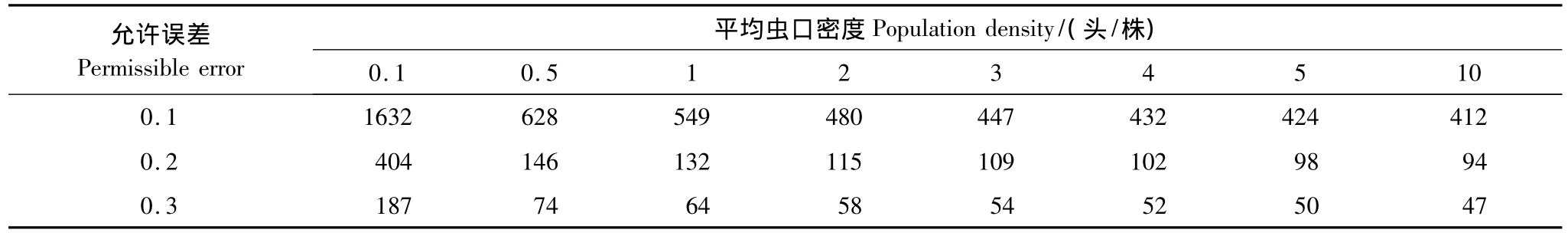
允许误差Permissible error平均虫口密度Population density/(头/株) 0.1 0.5 1 2 3 4 5 10 0.1 1632 628 549 480 447 432 424 412 0.2 404 146 132 115 109 102 98 94 0.3 187 74 64 58 54 52 50 47
通过对以上两种抽样方法比较可以看出:空间抽样可以根据不同的依赖范围,设定不同的抽样范围,抽样精度较高,基本都在80%以上。还能根据不同受害程度和林分类型分别单独进行抽样设计,具有很强的操作性和针对性;而传统抽样方法较单一,抽样结果与允许误差密切相关,抽样分析的结果常常是确定某种误差条件下的抽样数。
4 结论与讨论
受害程度不同的油松纯林林分内,红脂大小蠹种群空间格局具有一定的差异,混交林与纯林林分内其空间格局也具有较大的差别。混交林林分内红脂大小蠹表现为随机分布,而纯林为聚集分布。混交林林分内红脂大小蠹空间依赖范围远远大于纯林林分内的空间依赖范围。这说明红脂大小蠹空间格局,不仅与其各自的生物学特性有关,而且还与林分受害程度以及林分类型有关。国内外研究表明,寄主树木的挥发性物质是对红脂大小蠹起到干扰选择与聚集的主要化学物质[25-26],而混交林内其他树种有可能干扰了红脂大小蠹与油松的信息交流,从而影响红脂大小蠹在混交林内的空间分布状况。红脂大小蠹能通过松树挥发性物质中的3-(+)-蒈烯等单萜烯、红脂大小蠹本身产生的挥发性物质马鞭草烯酮的特定浓度来确定其适宜的寄主,其对寄主选择应该是综合各种因子,而远非对少数寄主挥发性成分的简单识别的过程[27-29],林分类型及其他环境条件的改变,也会对此产生影响[30]。
和传统的空间格局分析类似,地统计学研究种群空间格局的目的之一也是指导对生态学现象的空间抽样调查,抽样设计的好坏直接影响空间分析的结果。采用传统抽样方法对目标统计量进行精度计算时,由于将样本之间视为完全独立,并忽略部分因样本之间的相关性而产生的信息损失,从而使最终计算的估计误差要比实际值偏低,这种偏差受多种因素共同的影响,如地理对象空间相关性、抽样方法、样本数量、采样密度及样本点大小等[31]。空间抽样技术将抽样理论与害虫的生物学特性相结合,不仅能计算抽样的误差情况,而且还能探讨被研究对象的适当抽样数量与最适样方大小,这样就使研究质量有了可靠保证[19]。
地统计学与传统空间格局分析方法是相互补充的,综合运用这两类方法可更全面准确地了解所研究对象的空间规律。研究昆虫空间格局有助于了解昆虫的生态学特性,对于虫口数量调查取样、扩散范围的预测预报以及制订害虫防治策略等都具有重要的意义[20,32]。
由于调查年限较短,调查资料有限,应用地统计学对红脂大小蠹种群在时间上的依赖性与连续性的研究有待进一步完成。另外,在生态系统中,天敌、天敌与害虫、害虫间的种间种内效应非常复杂。因此进一步探讨以明确自然生态系统中各物种之间的空间格局关系很有必要。
致谢:感谢国家林业局山西太岳山森林生态站和游憩2006级史红萃、张玲同学在外业调查中的帮助。
References:
[1] Li J S,Chang G B,Song Y S,Wang Y W,Chang B S.Carrying out project management and controlling plague of red turpentine beetle.Forest Pest and Disease,2001,(4):41-44.
[2] Liu G S,Wang J H,Han H J,Wang J P.Occurrence and control of Dendroctonus valens.Plant Protection,2003,29(1):58-59.
[3] Smith R H.Red Turpentine Beetle.Forest Pest Leaflet 55.Washington DC:U.S.Department of Agriculture,1971:17-65.
[4] Song Y S,Yang A L,He N J.Pest risk analysis of red turpentine beetle.Forest Pest and Disease,2000,(6):34-37.
[5] Zhao J X,Yang Z Q,Ren X H,Liang X M.Biological characteristics and occurring law of Dendroctonus valens in China.Scientia Silvae Sinicae,2008,44(2):99-104.
[6] Ding Y Q.Entomological Mathematics Ecology.Beijing:Science Press,1994:22-69.
[7] Zheng H Y,Xia N B.Forestry Entomological Ecology.Beijing:China Forestry Publishing House,1993:120-167.
[8] Miao Z W.Space distribution of entering tree hole of Dendroctonus vales imago.Shanxi Forestry Science and Technology,2002,(3):7-9.
[9] Zhang L Y,Zhang J T,Zhai Y H.Chemical control strategies and practices for red turpentine beetle.Journal of Beijing Forestry University,2003,25(3):37-41.
[10] Li J K,Li Y Z,Li L,He H,Guo X R,Li M L.Distributional pattern and economic thresholds of Dendroctonus valens.Journal of Northwest Forestry University,2006,21(5):109-112.
[11] Li Y C,Xia N B,Tu H Q,Luo Y Q,Wen J B.A geostatistical analysis on spatial patterns of Anoplophora glabripennis in poplars.Acta Ecologica Sinica,1997,17(4):393-401.
[12] Shi G S,Li D M.Geostatistical analysis of spatial pattern of Dendrolimus punctatus.Chinese Journal of Applied Ecology,1997,8(6):612-616.
[13] Zhou G F,Xu R M.Biogeostatistics.Beijing:Science Press,1997:7-65.
[14] Wang Z Q.Application of Geostatistics in Ecology.Beijing:Science Press,1999:10-72.
[15] Yunsel T Y,Ersoy A,Cetin M.Geostatistical analysis of spatial distribution of salt bed thickness.Journal of Mining Science,2002,38(6):565-572.
[16] Hohn M E,Liebhold A M,Gribko L S.Geostatistical model for forecasting spatial dynamics of defoliation caused by the gypsy moth(Lepidoptera: Lymantriidae).Environmental Entomology,1993,22(5):1066-1075.
[17] Jia H M,Huang D Z.Research on the relationship between red turpentine and the type of forests.Journal of Anhui Agriculture Science,2006,34 (5):884-885.
[18] Zong S X,Luo Y Q,Xu Z C,Wang T,Kari H.Spatial distribution regularities of Holcocerus hippophaecolus pupae.Acta Ecologica Sinica,2006,26(10):3233-3237.
[19] Zhou G F,Xu R M.Biogeostatistics:Analysis of Spatio-temporal Dynamics of Biological Populations.Beijing:Science Press,1998:1-156.
[20] Chen H H,Zhang J W,Yang S L.Biogeostatistics and sampling technique about spatial pattern of Chinolyda flagellicornis diapause larva.Forest Research,2002,15(5):593-598.
[21] Shang S H,Zhang Y C.Study on spatial pattern of Liriomyza huidobrensis blanchard in tobacco field.Guizhou Agriculture Sciences,2002,30 (6):20-22.
[22] Ding Y Q.Theories and Applications of Entomological Mathematics Ecology.Beijing:Science Press,1980:18-31.
[23] Zhao D L.Forecast Diseases and Pests of Plant.Beijing:China Agriculture Publishing House,2000:40-46.
[24] Zahner R P.Sampling statistics for Panonychus nimi and Tetranychus urticae feeding on apple tree.Researches on Population Ecology,1984,(26):97-112.
[25] Vite J P,Gara R I.Volatile attractants from ponderosa pine attacked by bark beetles(Coleoptera:Scolytidae).Contributions from Boyce Thompson Institute,1962,(21):251-274.
[26] Owen D R.The Role of Dendroctonus valens and its Vectored Fungi in the Mortality of Ponderosa Pine.Berkeley(MI):University of California,1985.
[27] Zhang L W,Lu M,Liu Z D.Progress on invasion biology and chemical ecology of red turpentine beetle,Dendroctonus valens.Chinese Bulletin of Entomology,2007,44(2):172-177.
[28] Jia H M,Huang D Z,Cao Y X,Kong X B,Zhang Z.Relationship between resin monoterpenes and resistance of Chinese pine to red turpentine beetle.Journal of Northeast Forestry University,2008,36(1):48-50.
[29] Wang H B,Zhang Z,Kong X B.Relationship between release regularity of volatiles from Pinus tabulaeformis and the damage by Dendroctonus valens.Journal of Beijing Forestry University,2005,27(2):75-80.
[30] Wang H B,Wang Y G,Zhang Z,Jin Y J,Zhou S Z.The volatile components of the Chinese pine and the eag responses of the Dendroctonus valens (Coleoptera:Scolytidae).Scientia Silvae Sinicae,2006,42(5):85-88.
[31] Cao Z D,Wang J F,Li L F,Jiang C S.Strata efficiency and optimization strategy of stratified sampling on spatial population.Progress in Geography,2008,27(3):153-160.
[32] Farley R A,Fitter A H.Temporal and spatial variation in soil resources in a deciduous woodland.Journal of Ecology,1999,(87):668-696.
参考文献:
[1] 李计顺,常国彬,宋玉双,王艺伟,常宝山.实施工程治理控制红脂大小蠹虫灾——对红脂大小蠹暴发成灾及治理对策的探讨.中国森林病虫,2001,(4):41-44.
[2] 刘光生,王俊华,韩惠娟,王建平.红脂大小蠹发生危害及其防治.植物保护,2003,29(1):58-59.
[4] 宋玉双,杨安龙,何嫩江.森林有害生物红脂大小蠹的危险性分析.森林病虫通讯,2000,(6):34-37.
[5] 赵建兴,杨忠岐,任晓红,梁小明.红脂大小蠹的生物学特性及在我国的发生规律.林业科学,2008,44(2):99-104.
[6] 丁岩钦.昆虫数学生态学.北京:科学出版社,1994:22-69.
[7] 郑汉业,夏乃斌.森林昆虫生态学.北京:中国林业出版社,1993:120-167.
[8] 苗振旺.红脂大小蠹成虫侵入孔的空间分布型研究.山西林业科技,2002,(3):7-9.
[9] 张历燕,张京陶,翟银海.化学防治红脂大小蠹的策略与实践.北京林业大学报,2003,25(3):37-41.
[10] 李建康,李有忠,李莉,贺虹,郭新荣,李孟楼.红脂大小蠹的分布型与防治指标研究.西北林学院学报,2006,21(5):109-112.
[11] 李友常,夏乃斌,屠洪泉,骆有庆,温俊宝.杨树光肩星天牛种群空间格局的地统计学研究.生态学报,1997,17(4):393-401.
[12] 石根生,李典谟.马尾松毛虫空间格局的地学统计学分析.应用生态学报,1997,8(6):612-616.
[13] 周国法,徐汝梅.生物地理统计学.北京:科学出版社,1997:7-65.
[14] 王政权.地质统计学在生态学中的应用.北京:科学出版社,1999:10-72.
[17] 贾洪敏,黄大庄.灵空山红脂大小蠹发生与林分及其立地条件的关系.安徽农业科学,2006,34(5):884-885.
[18] 宗世祥,骆有庆,许志春,王涛,Kari Heliövaara.沙棘木蠹蛾蛹的空间分布.生态学报,2006,26(10):3233-3237.
[19] 周国法,徐汝梅.生物地理统计学:生物种群时空分析的方法及其应用.北京:科学出版社,1998:1-156.
[20] 陈绘画,张建薇,杨胜利.鞭角华扁叶蜂滞育幼虫空间格局的生物地理统计学分析及抽样技术的研究.林业科学研究,2002,15(5): 593-598.
[21] 商胜华,张永春.烟田南美斑潜蝇空间分布型研究.贵州农业科学,2002,30(6):20-22.
[22] 丁岩钦.昆虫数学生态学原理与应用.北京:科学出版社,1980:18-31.
[23] 赵德良.植物病虫测报.北京:中国农业出版社,2000:40-46.
[27] 张龙娃,鲁敏,刘柱东,孙江华.红脂大小蠹入侵机制与化学生态学研究.昆虫知识,2007,44(2):172-177.
[28] 贾洪敏,黄大庄,曹逸霞,孔祥波,张真.松脂单萜类物质与油松对红脂大小蠹抗性的关系.东北林业大学学报,2008,36(1):48-50.
[29] 王鸿斌,张真,孔祥波.油松萜烯类挥发物释放规律与红脂大小蠹危害的关系.北京林业大学学报,2005,27(2):75-80.
[30] 王鸿斌,王玉刚,张真,金幼菊,周淑芷.油松挥发性物质分析及红脂大小蠹的触角电位反应.林业科学,2006,42(5):85-88.
[31] 曹志冬,王劲峰,李连发,姜成晟.地理空间中不同分层抽样方式的分层效率与优化策略.地理科学进展,2008,27(3):153-160.
Geostatistical analysis and sampling technique on spatial distribution pattern of Dendroctonus valens population
基金项目:北京市教育委员会共建项目(JD100220888);中央高校基本科研业务费专项资金资助
收稿日期:2010-05-04;
修订日期:2010-10-25