
山西崦山自然保护区侧柏林植物生态位特征
【类型】期刊
【作者】赵天梁(山西省林业调查规划院)
【作者单位】山西省林业调查规划院
【刊名】北京林业大学学报
【关键词】 侧柏群落;优势种;生态位宽度;生态位重叠;崦山自然保护区
【资助项】山西省自然科学基金项目(2013011037-1);科技部科技基础性工作专项(2011fy110300)
【ISSN号】1000-1522
【页码】P24-30
【年份】2019
【期号】第8期
【期刊卷】1;|7;|8;|4;|5;|2
【摘要】应用Shannon-Weaver指数和Petraitis指数,研究了山西崦山自然保护区侧柏林主要植物的生态位(包括生态位宽度和生态位重叠)。结果表明:侧柏林36个优势种中,乔木中侧柏生态位宽度最大(3.56)、白皮松次之(3.12);灌木中荆条生态位宽度最大(3.43),黄刺玫(3.09)和多花胡枝子次之(3.09);草本中披针苔草和山蒿生态位宽度值较大(分别为3.39、3.03),主要是由于这些种具有较为广泛的适应性。生态位宽度与物种分布频数呈正相关(r=0.941,P<0.01),这说明物种的分布范围越广,它的生态适应性越强,因而生态位就越宽。36个优势种群并不存在完全普遍重叠,主要原因是环境的异质性较大和物种之间的生态互补性所致。在36个优势种中,表现出生态位重叠的有1 099个种对,占总种对数的87.22%。侧柏与大多数种的重叠值较小,这意味着与其他种相比,在对资源利用的竞争中侧柏处于相对有利的位置,对群落的适应性要优于其他植物。
【全文】 文献传递
山西崦山自然保护区侧柏林植物生态位特征
摘 要:应用Shannon-Weaver指数和Petraitis指数,研究了山西崦山自然保护区侧柏林主要植物的生态位(包括生态位宽度和生态位重叠)。结果表明:侧柏林36个优势种中,乔木中侧柏生态位宽度最大(3.56)、白皮松次之(3.12);灌木中荆条生态位宽度最大(3.43),黄刺玫(3.09)和多花胡枝子次之(3.09);草本中披针苔草和山蒿生态位宽度值较大(分别为3.39、3.03),主要是由于这些种具有较为广泛的适应性。生态位宽度与物种分布频数呈正相关(r=0.941,P<0.01),这说明物种的分布范围越广,它的生态适应性越强,因而生态位就越宽。36个优势种群并不存在完全普遍重叠,主要原因是环境的异质性较大和物种之间的生态互补性所致。在36个优势种中,表现出生态位重叠的有1 099个种对,占总种对数的87.22%。侧柏与大多数种的重叠值较小,这意味着与其他种相比,在对资源利用的竞争中侧柏处于相对有利的位置,对群落的适应性要优于其他植物。
关键词:侧柏群落;优势种;生态位宽度;生态位重叠;崦山自然保护区
生态位研究是种群生态学的重要研究内容之一。生态位研究结果可以帮助人们从理论上全面了解种群的生态适应性、种间竞争和共存的机理,有助于理解种群在资源共享和利用中的地位角色和作用,有助于揭示种群在群落组成和结构以及在生态系统中的功能地位[1-4]。自生态位理论提出后,许多学者提出了各种生态位测度[5-11],这些方法作为种群生态学的重要分析手段得到广泛的应用,如Pianka[6]关于蜥蜴(Eumeces fasciatus)群落生态位的研究;Anthwal等[12]关于印度喜马拉雅山区草地植被结构、生态位宽度和重叠以及竞争关系的研究;王孝安[13]关于马衔山优势种群的竞争研究;郑元润[14]关于大青沟森林群落主要木本植物生态位的研究;张峰等[1]关于翅果油树(Elaeagnusmollis)群落优势种群生态位的分析;邓永利等[15]关于山西万家寨引黄工程北干线沿线植被优势种群生态位研究。
山西崦山自然保护区是山西省侧柏(Platycladus orientalis)林分布最集中的地区之一,群落结构完整,林相整齐,具有山西温性针叶林的典型特征[12]。本文采用Shannon-Weaver指数Petraitiss方法研究了山西崦山自然保护区侧柏林优势种的生态位宽度及生态位重叠,目的是揭示侧柏林优势种生态位宽度与生态适应性的关系,物种分布频数与生态位的关系以及种间生态位重叠与植物生活型、群落稳定性的关系,旨在为崦山自然保护区侧柏资源的科学管理和保护提供理论基础。
1 研究区域自然地理概况
山西崦山自然保护区位于中条山东段,行政区划属于山西省阳城县,地理位置:35°35′33″~35°41′41″N, 112°16′50″~112°27′50″E,海拔600~1 151m。总面积8 992.6 hm2。其他自然地理因素见文献[16]。
2 研究方法
2.1 野外取样
调查参照方精云等[17]方法,并结合侧柏林分布生境异质性较小和群落组成结构较为简单等实际情况稍作修改。样方面积10m×10m,野外共记录37个样方。记录的群落信息包括:1)群落总盖度、乔木层盖度,树高、胸径、冠幅及每种乔木的盖度;灌木层样方面积4m×4m,记录的信息包括灌木层分盖度、每种灌木的高度、盖度和丛数;草本层样方面积1m×1 m,记录草本分层盖度、每种草本植物的高度、盖度等。2)记录的环境信息包括海拔、坡度、坡向、人为干扰情况和经纬度。共记录了60种种子植物。
2.2 生态位分析
根据植物在侧柏群落中的作用和地位,选择36个优势种(表1)进行生态位宽度和种间生态位重叠的测定。
2.2.1 生态位宽度
采用Shannon-Weaver指数,分别以样方和坡向作为资源状态,以每个种的重要值来测定物种的生态位宽度[1,18]:
式中:nij为第i个种在第j个资源轴的重要值,Nij为第j个资源轴上所有种的重要值之和;r为资源轴的数目。
应用Pearson相关系数分析物种分布频数(f)与生态位宽度的相关性。
2.2.2 生态位重叠
2.2.2.1 生态位特定重叠
按照Petraitis[8-9]关于种间生态位重叠的定义, 种i与种j的特定重叠是不对称的,即种i与种j的重叠指数不等于种j与种i的重叠指数SOij计算公式[1-2,18]。
种i与种j的特定重叠指数[1,8-9]为:

式中:Pik与Pjk分别为第i个种和第j个种的重要值在第k个资源轴所占的比例,nik与njk分别为第i个种和第j个种在第k个资源轴上的重要值,cj为第j个资源轴的重要值之和占总重要值的比例,tj和tk分别为第j个轴和第k轴的所有种的重要值之和,T所有种的重要值之和,s为总种数。
特定重叠指数可以用χ2检验来判断其显著性,对应的统计量分别是:Uij=-2Niln(SOij)和Uji= -2Njln(SOji)。如果Uij>χ20.05,则种i和j之间不存在特定重叠;如果Uij<χ20.05,则种i和j之间存在特定重叠。
2.2.2.2 生态位普遍重叠的测定
生态位普遍重叠指数GO[1,8-9,18]公式为:
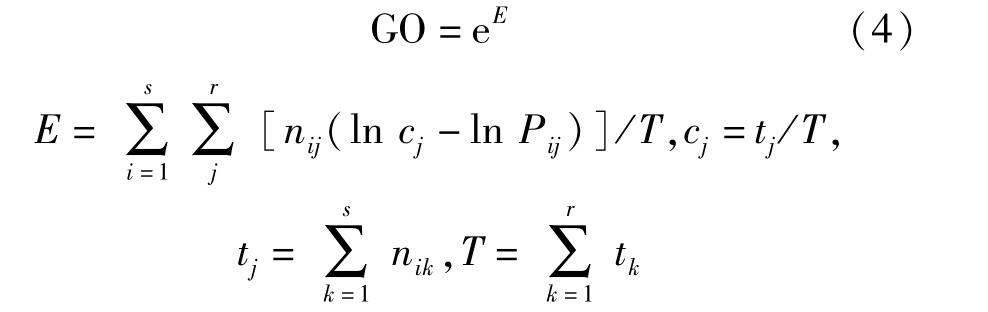
与特定重叠指数类似,普遍重叠指数也可以进行χ2显著性检验,统计量为V=-2Tln(GO)。V>χ20.05种i和j之间完全普遍重叠不存在;V<χ20.05,种i和j之间完全普遍重叠存在。
3 结果与分析
3.1 生态位宽度分析
生态位宽度不仅与物种本身的生理和生态适应性、竞争力、种间竞争强度密切相关,而且与可利用资源数量和分布空间依存度极大。如果物种在群落和生态系统中扮演着主要的角色,处于优势或支配地位[1],就可能控制着群落的演替方向和稳定性,很大程度上决定了生态系统物质循环和能量流动的方式和过程。
侧柏林优势种的生态位宽度测定结果见表1。
表1 侧柏群落优势种群生态位宽度
Tab.1 Niche breadth of dominant populations inPlatycladus orientalisforest

序号Species No.种名Species频数Frequency (f)生态位宽度Niche breadth(B)序号Species No.种名Species频数Frequency (f)生态位宽度Niche breadth(B) 1侧柏Platycladus orientalis37 3.56 19山蒿Artemisia brachyloba25 3.03 2白皮松Pinusbungeana25 3.12 20米口袋Gueldenstaedtia multiflora16 2.63 3栓皮栎Quercus variabilis5 1.58 21白羊草Bothriochloa ischemum18 2.57 4辽东栎Q.wutaishannica3 1.03 22细叶远志Polygala tenuifolia9 2.07 5黄连木Pistacia chinensis2 0.51 23委陵菜Potentilla chinensis6 1.75 6荆条Vitex negundovar.heterophylla36 3.43 24斑叶堇菜Viola variegata7 1.74 7黄刺玫Rosa xanthina26 3.09 25早开堇菜Viola prionantha6 1.58 8多花胡枝子Lespedeza floribunda28 3.09 26雅葱Scorzonera austriaca5 1.58 9白刺花Sophora viciifolia19 2.83 27草地风毛菊Saussurea amara4 1.3 10少脉雀梅藤Sageretia paucicostata16 2.56 28冰草Agropyron cristatum4 1.30 11达乌里胡枝子Lespedeza davurica10 2.14 29荩草Arthraxon hispidus4 1.13 12三裂绣线菊Spiraea trilobata9 1.95 30石沙参Adenophora polyantha3 1.07 13毛黄栌Cotinus coggygriavar.pubescens5 1.57 31地梢瓜Cynanchum thesioides3 1.02 14灰栒子Cotoneaster acutifolius5 1.54 32火绒草Leontopodium leontopodioides3 0.95 15美丽胡枝子Lespedeza formosa5 1.49 33双花堇菜Viola biflora4 1.35 16陕西荚蒾Viburnum schensianum5 1.32 34黄背草Themeda japonica3 0.66 17小叶鼠李Rhamnusparvifolia4 1.19 35鼠麴草Gnaphalium affine2 0.67 18披针苔草Carex lanceolata32 3.39 36狗尾草Setaria viridis2 0.25
从表1可以看出,侧柏生态位宽度最大(3.56),这是因为侧柏广泛分布于调查的所有样地,作为建群种高居于群落最上层,能够最大限度地利用光资源;并且在所有乔木中侧柏种群密度最大(6.28±0.520)株/hm2,DBH最大(20.56±1.13) cm,表明侧柏对崦山保护区的生态环境具有广泛的适应性,对资源的综合利用能力最强,且竞争能力较强。白皮松(Pinus bungeana)是乔木层的共群种或优势种,分布较广(频数f=24),DBH次之(15.54± 2.14)cm,因此具有较大的生态位宽度。栓皮栎(Quercus variabilis)、辽东栎(Q.wutaishannica)和黄连木(Pistacia chinensis)出现频数(f=5、3和2)较低,仅在水分条件较好的阴坡和半阴坡出现,这说明它们分布的生态环境有局限性,一定程度上限制了它们对生态环境的适应性和对资源的利用能力,因此生态位宽度较小。
灌木的生态位宽度依次为:荆条(Vitex negundovar.heterophylla)>黄刺玫(Rosa xanthina)>多花胡枝子(Lespedeza floribunda)>少脉雀梅藤(Sageretia paucicostata)>白刺花(Sophora viciifolia)>达乌里胡枝子(Lespedeza.davurica)>毛黄栌(Cotinus coggygriavar.pubescens)>陕西荚蒾(Viburnum schensianum)>灰栒子(Cotoneaster acutifolius)>小叶鼠李(Rhamnus parvifolia)。荆条和黄刺玫频数最高(f分别为36和26),作为多年生中生植物具有广泛的适应性,位于灌木层的第一亚层,不仅能够在光照充足、土壤水分条件较差的阳坡广泛分布,也能在立地条件较好的阴坡生存和繁衍,因此表现较大的生态位宽度。位于灌木层的第二亚层的多花胡枝子,其空间生态位与其他种实现了分化,从而一定程度上避免了与其他种对空间资源的竞争,并能充分利用下层空间与其他种实现共存,因此生态位也较宽。毛黄栌、陕西荚蒾和灰栒子在群落中的数量较少(f=5),在空间分布范围较窄,大大限制了对生长发育所需资源(包括水分、养分等)的利用能力,因此它们具有较小的生态位宽度。
草本植物中披针苔草(Carex lanceolata)、山蒿(Artemisia brachyloba)和白羊草(Bothriochloa ischemum)为草本层的优势种,也是山西暖温带落叶阔叶林地带草本植物群落的建群种;同时也是森林、灌丛、草本植物群落的优势种或常见种,具有分布区域大、生理生态适应性强的特点,既能在水土流失的黄土丘陵沟壑区分布,也能在低中山山地生存和繁衍,表现出广泛的生态和生理适应性和较大的生态位宽度。狗尾草(Setaria viridis)在山西广泛分布于农田和果园等人工植物群落,为杂草,具有极强的适应性,但仅在样方13和样方24存在,故其生态位宽度最小(仅为0.25)。
3.2 物种分布频数(f)与生态位宽度关系分析
物种分布频数(f)与生态位宽度关系见图1。物种分布频数与生态位宽度的相关系数(r=0.941,P<0.01)表明:物种频数与生态位宽度为极显著的正相关,说明一个种的分布频数越大,其生态位宽度就越大。如频数最大(f=36)的侧柏,其生态位宽度最大;而频数最小(f=2)的狗尾草,其生态位宽度最小。这是因为一个种的分布频数越大,意味着其分布范围就越大,物种对异质性的生境适应的生态幅就越大,因而使得它有较大的生态位宽度。
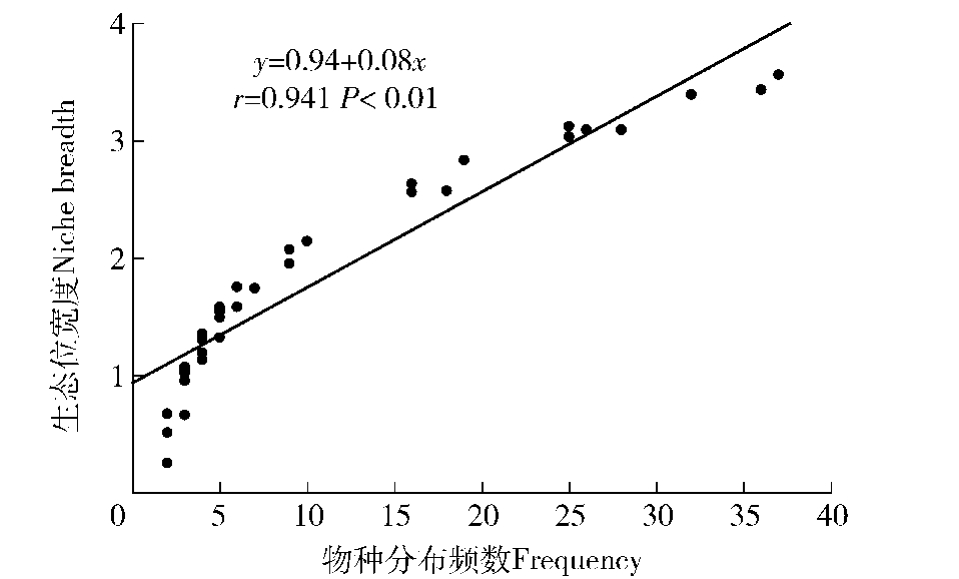
图1 侧柏林优势种群生态位宽度与频数的关系
Fig.1 Relationship between frequency and niche breadth of dominant populations in P.orientalis forest
3.3 生态位重叠
3.3.1 生态位重叠测定结果
侧柏林生态位普遍重叠指数:GO=0.522(V= 14 074.63,P<0.01),这意味着36个种完全普遍重叠并不存在。主要是由于:1)生境多样性。侧柏林分布的区域既有生境条件较好的阴坡,也有生境条件较差、基岩裸露的严重退化区域,生境的差异性会限制某些种的分布,如基岩裸露的生境仅适于耐旱性较强的植物分布,不适于对水分需求较大的种分布,因而使得不同区域侧柏群落的结构和物种组分产生明显分化,使得所有普遍重叠指数不显著。2)物种对生境适应性的分化。由于不同植物对各种资源(如光照、水分、土壤养分等)需求不同,这将促使生态位在进化过程中产生明显的分化,最终导致种间生态位重叠程度大大降低。如位于侧柏林下的荆条、黄刺玫、少脉雀梅藤、白刺花、白羊草和山蒿等为阳性植物,而灰栒子、陕西荚蒾、草地风毛菊(Saussurea amara)、双花堇菜(Viola biflora)等相对较为耐荫,它们生态和生物学上的这种互补性,使得这些种之间最大限度地避免了对资源的竞争,特别是对于光资源的竞争,从而使得它们之间的生态位重叠值较小。
3.3.2 生态位特定重叠测定结果
侧柏林植物种间生态位重叠测定结果表明(表2),36种植物构成了1 260个种对,其中有1 099个种对表现出生态位重叠,占总数的87.22%,这意味着36种植物所利用的资源类型相似度较高。侧柏与其余物种的生态位重叠值较小,这说明侧柏与其他种在资源利用方面的竞争力较强,对群落组分、结构、功能具有决定性的作用,对维系侧柏林的稳定至关重要。
一般来说,生态位重叠值较大的种对预示着这两个种可能具有相似的生态学和生物学特性,或者对资源的需求有一定的互补性[13,18-19]。如栓皮栎与荆条同为群落的优势种尽管生态位重叠值较大,但栓皮栎位于乔木层,荆条位于灌木层,占据不同的高度,空间生态位出现明显的分异现象,对生存空间形成明显的互补关系,因此它们会实现相对稳定的共存。
3.3.3 物种生活型与生态位重叠的测定结果
种间生态位重叠与物种生活型有一定关联。表3是侧柏林不同生活型物种与种间生态位重叠比例的关系,可以看出:在相同生活型的植物中乔木之间的生态位重叠比例(85.00%)最低,且重叠值普遍较小;如侧柏与白皮松、黄连木、栓皮栎、辽东栎的重叠值最大的仅为0.032 3(侧柏与黄连木),最小的仅为0.007(侧柏与白皮松)。灌木之间的生态位重叠比例不仅最高(高达99.24%),而且生态位重叠值也较大,如黄刺玫除了与荆条和陕西荚蒾外,与少脉雀梅藤、白刺花、毛黄栌的重叠值都较高。这是由于灌木种群密度较大,导致它们的灌木层物种呈镶嵌分布,从而使得灌木之间生态位重叠的种对较多。
表2 侧柏群落优势种生态位特定重叠测定结果
Tab.2 Specific niche overlapping of dominant species in P.orientalis forest

注:*P<0.05;**P<0.01;序号同表1。No.of species is the same as that in table 1.
种Species 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 2425 26 27 28 29 30 31 32 33 34 35 36 1 0.001 0.032 0.006 0.005 0.021 0.002 0.000 0.000 0.000 0.002 0.001 0.000 0.055 0.004 0.010 0.009 0.001 0.000 0.001 0.001 0.000 0.000 0.005 0.000 0.004 0.007 0.001 0.000 0.002 0.000 0.0000.000 0.000 0.000 0.000 2 0.057 0.322*0.005 0.002 0.023 0.007 0.035 0.006 0.004 0.001 0.003 0.002 0.013 0.001 0.004 0.002 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 3 0.004 0.005 0.007 0.003 0.018 0.001 0.004 0.000 0.001 0.016 0.003 0.001 0.015 0.001 0.001 0.003 0.000 0.000 0.007 0.001 0.000 0.000 0.000 0.000 0.002 0.001 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 4 0.003 0.000 0.001 0.002 0.434**0.007 0.001 0.002 0.000 0.000 0.040 0.006 0.007 0.065 0.002 0.039 0.001 0.000 0.001 0.006 0.133 0.000 0.000 0.000 0.001 0.018 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 5 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.000 0.000 0.001 0.000 0.000 0.001 0.000 0.000 0.000 0.032 0.000 0.010 0.025 0.000 0.003 0.000 0.000 0.000 0.019 0.002 0.000 0.000 0.000 0.002 0.000 0.000 0.0001 6 0.001 0.000 0.000 0.003 0.001 0.008 0.002 0.000 0.000 0.001 0.032 0.008 0.002 0.117 0.001 0.019 0.001 0.000 0.001 0.017 0.048 0.000 0.000 0.000 0.001 0.004 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 7 0.000 0.000 0.000 0.000 0.023 0.001 0.002 0.004 0.000 0.000 0.002 0.000 0.007 0.002 0.000 0.000 0.000 0.000 0.001 0.005 0.008 0.000 0.000 0.000 0.000 0.000 0.002 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 8 0.012 0.006 0.022 0.002 0.016 0.376*0.089 0.176 0.008 0.011 0.051 0.004 0.008 0.206 0.014 0.034 0.002 0.000 0.001 0.016 0.024 0.000 0.001 0.000 0.000 0.015 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 9 0.000 0.008 0.000 0.002 0.057 0.269 0.100 0.112 0.103 0.001 0.202 0.017 0.013 0.146 0.001 0.031 0.004 0.066 0.014 0.060 0.104 0.002 0.000 0.010 0.010 0.002 0.009 0.014 0.001 0.001 0.001 0.002 0.001 0.000 0.000 10 0.001 0.016 0.001 0.012 0.026 0.215 0.123 0.086 0.358*0.004 0.545**0.014 0.190 0.121 0.008 0.107 0.001 0.000 0.009 0.037 0.104 0.001 0.000 0.001 0.016 0.009 0.000 0.000 0.002 0.000 0.001 0.000 0.001 0.012 0.000 11 0.001 0.000 0.014 0.000 0.000 0.205 0.000 0.002 0.000 0.000 0.083 0.134 0.000 0.060 0.003 0.006 0.002 0.000 0.000 0.026 0.029 0.001 0.003 0.000 0.002 0.042 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.002 0.000 12 0.002 0.015 0.001 0.004 0.057 0.362*0.091 0.035 0.130 0.044 0.005 0.022 0.084 0.191 0.003 0.069 0.001 0.000 0.016 0.060 0.046 0.001 0.000 0.001 0.008 0.003 0.000 0.000 0.001 0.000 0.000 0.000 0.000 0.002 0.000 13 0.000 0.001 0.000 0.003 0.000 0.344*0.006 0.001 0.002 0.000 0.011 0.076 0.002 0.090 0.002 0.015 0.000 0.000 0.004 0.007 0.037 0.022 0.000 0.000 0.001 0.019 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 14 0.015 0.000 0.001 0.022 0.078 0.244 0.036 0.002 0.003 0.006 0.004 0.366*0.015 0.337*0.008 0.134 0.002 0.000 0.016 0.050 0.126 0.001 0.001 0.002 0.018 0.014 0.007 0.000 0.003 0.000 0.000 0.000 0.000 0.040 0.000 15 0.001 0.000 0.012 0.001 0.021 0.298 0.025 0.001 0.001 0.000 0.001 0.029 0.003 0.014 0.009 0.039 0.004 0.000 0.001 0.005 0.049 0.000 0.000 0.000 0.001 0.020 0.002 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 16 0.367*0.001 0.035 0.015 0.014 0.213 0.033 0.001 0.001 0.001 0.012 0.057 0.003 0.187 0.694**0.200 0.010 0.000 0.004 0.016 0.155 0.000 0.004 0.002 0.015 0.111 0.004 0.000 0.006 0.001 0.001 0.000 0.000 0.017 0.000 17 0.020 0.000 0.036 0.0128 0.027 0.251 0.009 0.001 0.001 0.001 0.010 0.081 0.002 0.081 0.301 0.022 0.225 0.093 0.023 0.109 0.597 0.001 0.000 0.025 0.247 0.113 0.032 0.085 0.022 0.000 0.001 0.010 0.000 0.001 0.020 18 0.024 0.000 0.040 0.0095 0.062 0.289 0.017 0.001 0.001 0.001 0.014 0.062 0.002 0.159 0.647**0.029 0.618 0.330 0.027 0.231 0.529 0.001 0.007 0.027 0.248 0.100 0.504 0.253 0.130 0.001 0.004 0.044 0.000 0.007 0.111 19 0.000 0.000 0.000 0.0008 0.001 0.001 0.014 0.000 0.000 0.000 0.000 0.000 0.000 0.001 0.001 0.000 0.000 0.001 0.003 0.007 0.098 0.000 0.000 0.001 0.006 0.001 0.294 0.227 0.002 0.000 0.000 0.003 0.000 0.000 0.003 20 0.000 0.000 0.009 0.0002 0.090 0.001 0.032 0.000 0.000 0.000 0.006 0.011 0.003 0.040 0.003 0.001 0.001 0.004 0.206 0.137 0.019 0.002 0.002 0.102 0.032 0.006 0.349 0.022 0.009 0.002 0.017 0.019 0.008 0.004 0.101 21 0.003 0.000 0.001 0.002 0.036 0.410**0.013 0.003 0.005 0.002 0.002 0.108 0.005 0.058 0.141 0.003 0.071 0.017 0.120 0.019 0.155 0.001 0.008 0.019 0.146 0.003 0.041 0.020 0.034 0.000 0.001 0.029 0.000 0.000 0.021 22 0.000 0.000 0.000 0.004 0.001 0.122 0.000 0.000 0.000 0.000 0.000 0.009 0.001 0.000 0.047 0.001 0.009 0.004 0.029 0.000 0.011 0.000 0.000 0.000 0.004 0.011 0.003 0.015 0.000 0.000 0.000 0.0030 0.000 0.000 0.000 23 0.000 0.000 0.000 0.004 0.001 0.158 0.000 0.000 0.001 0.000 0.004 0.075 0.012 0.001 0.028 0.001 0.013 0.005 0.025 0.001 0.098 0.511 0.000 0.001 0.041 0.006 0.007 0.011 0.002 0.000 0.001 0.017 0.001 0.001 0.014 24 0.000 0.000 0.000 0.000 0.032 0.001 0.001 0.000 0.000 0.000 0.000 0.002 0.001 0.000 0.001 0.000 0.000 0.000 0.000 0.000 0.006 0.004 0.000 0.001 0.000 0.000 0.001 0.000 0.000 0.001 0.000 0.000 0.000 0.005 0.000 25 0.000 0.000 0.000 0.002 0.039 0.001 0.008 0.000 0.000 0.000 0.000 0.006 0.000 0.016 0.001 0.000 0.001 0.001 0.202 0.088 0.040 0.077 0.001 0.003 0.066 0.001 0.046 0.062 0.033 0.023 0.007 0.004 0.026 0.002 0.070 26 0.008 0.000 0.000 0.001 0.001 0.193 0.008 0.000 0.001 0.001 0.001 0.036 0.002 0.016 0.064 0.003 0.128 0.014 0.221 0.007 0.088 0.534 0.001 0.000 0.002 0.023 0.056 0.204 0.034 0.000 0.002 0.019 0.001 0.003 0.038 27 0.003 0.000 0.012 0.012 0.000 0.247 0.005 0.000 0.001 0.000 0.025 0.016 0.040 0.002 0.148 0.007 0.057 0.002 0.000 0.001 0.003 0.208 0.000 0.000 0.000 0.002 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 28 0.000 0.0000 0.000 0.001 0.001 0.001 0.020 0.000 0.000 0.000 0.000 0.002 0.000 0.018 0.001 0.000 0.001 0.001 0.545**0.010 0.089 0.117 0.000 0.000 0.002 0.081 0.001 0.109 0.024 0.001 0.003 0.014 0.000 0.002 0.171 29 0.001 0.000 0.000 0.007 0.001 0.116 0.016 0.000 0.001 0.000 0.001 0.001 0.016 0.006 0.133 0.002 0.041 0.013 0.732**0.003 0.014 0.528 0.000 0.000 0.001 0.033 0.047 0.245 0.005 0.001 0.001 0.015 0.000 0.000 0.005 30 0.010 0.000 0.000 0.015 0.001 0.199 0.012 0.000 0.001 0.001 0.001 0.027 0.002 0.041 0.124 0.008 0.101 0.034 0.377*0.009 0.138 0.433 0.001 0.015 0.010 0.308 0.012 0.574**0.282 0.190 0.084 0.033 0.007 0.019 0.316 31 0.000 0.000 0.000 0.000 0.001 0.000 0.011 0.000 0.000 0.000 0.000 0.002 0.001 0.002 0.001 0.000 0.000 0.000 0.189 0.004 0.020 0.017 0.001 0.002 0.017 0.035 0.001 0.318 0.036 0.030 0.178 0.012 0.038 0.005 0.3206 32 0.002 0.005 0.001 0.001 0.001 0.106 0.016 0.000 0.001 0.002 0.006 0.064 0.082 0.013 0.102 0.007 0.029 0.013 0.226 0.051 0.058 0.068 0.003 0.003 0.017 0.115 0.006 0.368*0.046 0.073 0.561**0.064 0.057 0.023 0.328 33 0.000 0.000 0.000 0.003 0.001 0.105 0.001 0.000 0.001 0.000 0.004 0.023 0.023 0.001 0.070 0.002 0.005 0.010 0.078 0.000 0.121 0.105 0.001 0.000 0.000 0.024 0.004 0.075 0.008 0.002 0.000 0.004 0.000 0.003 0.101 34 0.005 0.001 0.001 0.013 0.001 0.088 0.007 0.000 0.001 0.004 0.001 0.032 0.006 0.029 0.044 0.007 0.044 0.013 0.289 0.058 0.031 0.123 0.004 0.000 0.025 0.293 0.009 0.384 0.163 0.172 0.355*0.226 0.021 0.166 0.201 35 0.002 0.000 0.000 0.011 0.002 0.100 0.000 0.000 0.000 0.002 0.009 0.069 0.044 0.001 0.075 0.002 0.010 0.001 0.000 0.000 0.005 0.139 0.001 0.002 0.000 0.004 0.026 0.001 0.000 0.001 0.001 0.001 0.000 0.001 0.000 36 0.000 0.000 0.000 0.001 0.001 0.000 0.007 0.000 0.000 0.000 0.000 0.001 0.000 0.006 0.001 0.000 0.001 0.001 0.364 0.009 0.024 0.077 0.000 0.001 0.007 0.062 0.003 0.555**0.179 0.044 0.042 0.007 0.017 0.002 0.007
不同生活型之间生态位重叠比例的顺序为:乔木与灌木的生态位重叠比例(90.00%)>灌木层与草本层之间的重叠比例(83.55%)>乔木层与草本层之间重叠比例(76.32%)。
4 讨 论
4.1 生态位普遍重叠测定方法的比较
常用的生态位重叠测度中应用较为广泛的当属Pianka指数和Patraitis指数。绝大多数生态位重叠测度都假设两个种的重叠对称,优点是计算简单、快捷,但缺点是忽略了不同生活型种间竞争的不对称性。如侧柏林中乔木与灌木和草本植物的竞争就属于这种情况,侧柏在与其他种的竞争中,不仅地上部分,而且地下部分都处于绝对优势,即侧柏对灌木和草本植物的竞争力要高于灌木和草本植物对侧柏的竞争力。
表3 侧柏林优势种群生态位重叠与生活型的关系
Tab.3 Relationship between life form and niche overlapping of dominant populationsin P.orientalis forest
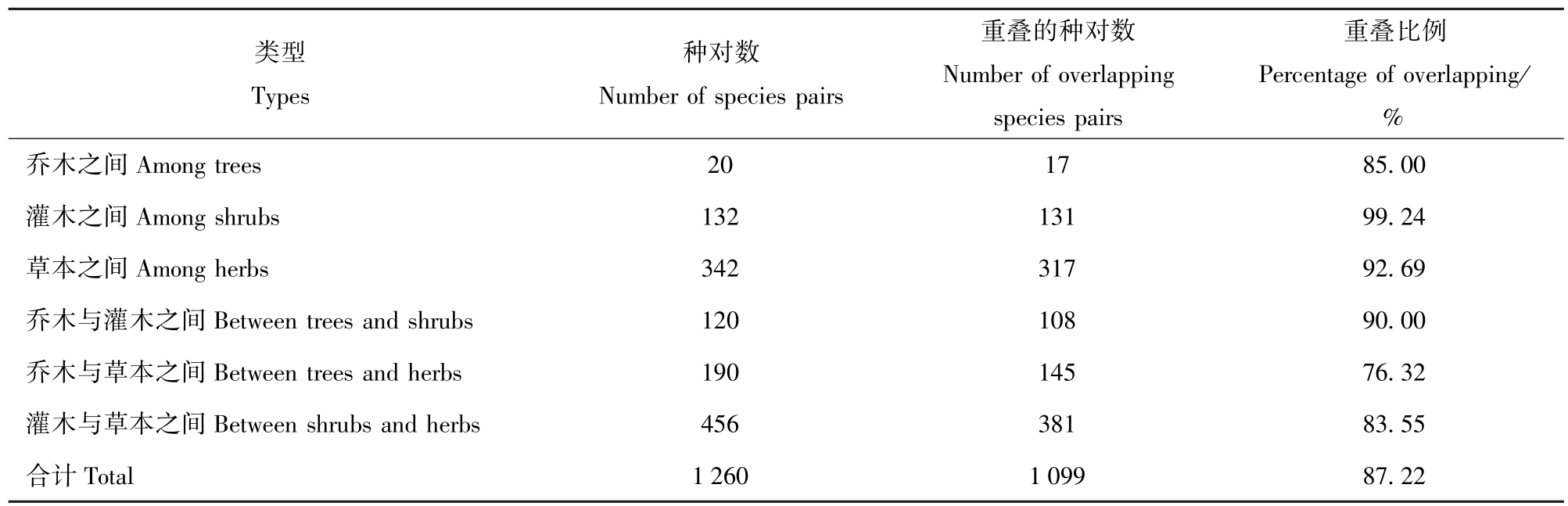
类型Types种对数Number of species pairs重叠的种对数Number of overlapping species pairs重叠比例Percentage of overlapping/%乔木之间Among trees 20 17 85.00灌木之间Among shrubs 132 131 99.24草本之间Among herbs 342 317 92.69乔木与灌木之间Between trees and shrubs 120 108 90.00乔木与草本之间Between trees and herbs 190 145 76.32灌木与草本之间Between shrubs and herbs 456 381 83.55合计Total 1 260 1 099 87.22
Patraitis指数计算生态位重叠时,考虑了种间竞争的不对称性,因此种i和种j的重叠值与种j和种i的重叠值并不相等。此外,与其他生态位重叠指数相比,Patraitis指数的测定结果还可以进行显著性检验,从而保证生态位重叠的测度具有统计学意义。
4.2 物种生活型与生态位重叠的关系
从表3可以看出:生活型相同的物种之间生态位重叠的比例要明显高于不同生活型植物间的比例。这是由于生活型相同的植物具有较为相似的生理和生态适应对策(如相似的光饱和点和光补偿点、相似的水分需求、相似的空间分布类型等),如乔木都位于侧柏林上层,因而导致它们之间的生态位重叠值较大[21]。而不同生活型的物种则不然,比如草本层的大多数植物光饱和点和光补偿点就要比乔木低,如披针苔草、斑叶堇菜(Viola variegata)、双花堇菜等,因此草本植物与木本植物(乔木和灌木)的生态位重叠对数就较低。
4.3 生态位重叠与种间竞争和群落稳定性的关系
生态位重叠分析结果表明:崦山自然保护区若干生活型相同的种间生态位重叠值较大的,如荆条分别与少脉雀梅藤、三裂绣线菊(Spiraea trilobata)、白刺花和毛黄栌,多花胡枝子与达乌里胡枝子等。主要是因为这些种对的生活型相同,生态习性、分布的生境较为相似所致。此外,一些不同生活型种对间的生态位重叠值也较大,如侧柏与小叶鼠李、荆条与火绒草(Leontopodium leontopodioides)、灰栒子与披针苔草等;由于这些种的生活型不同,在侧柏林中的层片亦不同,实现了空间生态位的分化。因此,空间的生态位重叠只发生在水平维度,这就很大程度上避免了种间的剧烈竞争[20],有助于侧柏林群落的相对稳定。
进一步分析侧柏群落微生境和各优势种的生态学和生物学特性分化程度,就会发现若干的种间生态位可能是完全或部分分离,如乔木层与草本层植物光饱和点和补偿点的分化,就会使种间生态位发生分离,从而对光资源的竞争强度大大降低。乔木拥有发达的根系,可以充分利用深层土壤深处的资源;而草本植物根系较浅,大多数利用土壤的表层资源,一定程度上可以避免它们对土壤资源的剧烈竞争和排斥作用的发生,从而有助于这些种长期共存和协同进化,维持侧柏林生态系统组分、结构、功能的相对稳定性。
5 结 论
1)山西崦山自然保护区侧柏林乔木层中生态位宽度占优势的为侧柏(生态位宽度3.56,下同)和白皮松(3.12)等;灌木层植物生态位宽度占优势的有荆条(3.43)、黄刺玫(3.09)和多花胡枝子(3.09) 等;草本层植物生态位宽度占优势的为披针苔草(3.39)、山蒿(3.03)等。
2)侧柏林36种植物构成的1 260个种对中有1 099个种对间存在生态位重叠,占总数的87.22%。
3)生活型相同的种间生态位重叠的比例要明显高于不同生活型植物间的比例。
参考文献:
[1]张峰,上官铁梁.翅果油树群落优势种群生态位分析[J].西北植物学报,2004,24(1):70-74.
ZHANG F,SHANG GUAN T L.Niche characteristics of dominant populations inElaeagnus molliscommunities,Shanxi[J].Acta Botanica Boreali-Occidentalia Sinica,2004,24(1):70-74.
[2]SYLVAIN D,DANIEL C,CLEMENTINE G C.Niche separation in community analysis:a new method[J].Ecology,2000,81:2914-2927.
[3]MUSTSHINDA C M,O'HARA R B.Integrating the niche and neutral perspectives on community structure and dynamics[J]. Oecologia,2011,166:241-251.
[4]THUILLERW,GASSO′N,PINA J,et al.Ecological niche and species traits:key drivers of regional plant invader assemblages [J].Biol Invasions,2012,14:1963-1980.
[5]COLWELL R K,FYTUYAMA D J.On themeasurementof niche breadth and overlap[J].Ecology,1971,52:567-576.
[6]PIANKA E R.The structure of lizard communities[J].Annual Review of Ecology and Systematics,1973,4:53-74.
[7]HANSHY I.Some comments on themeasurement of nichemetrics [J].Ecology,1978,59:168-174.
[8]PETRAIRTISPS.Likelihood measurements of niche breadth and overlap[J].Ecology,1979,60:703-710.
[9]PETRAIRTIS P S.The relationship between likelihood niche measurements and replicated tests for goodness of fit[J].Ecology, 1985,66:1983-1985.
[10]ABRAMSR.Some comments on measuring niche overlap[J]. Ecology,1980,61:44-49.
[11]李德志,石强,臧润国,等.物种或种群生态位宽度与生态位重叠的计测模型[J].林业科学,2006,42(7):95-103.
LID Z,SHIQ.ZANG R G,et al.Models for niche breadth and niche overlap of species or population[J].Scientia Silvae Sinica, 2006,42(7):95-103.
[12]ANTHWAL S,AJAY B,BHATT A B,et al.Vegetation structure,niche width,niche overlap and types of competition in temperate grazingland of Garhwal Himalaya,India[J]. Environmentalist,2008,28:261-273.
[13]王孝安.马衔山林区优势植物种群竞争的初步研究[J].植物生态学与地植物学丛刊,1984,8(1):36-40.
WANG X A.Preliminary study on the competiton among dominant plant populations in the Mahan Mountais forest zone[J].Acta Phytoecologica et Geobotanica,1984,8(1):36-40.
[14]郑元润.大青沟森林植物群落主要木本植物生态位研究[J].植物生态学报,1999,23(5):475-479.
ZHENG Y R.Main woody species niche of plant community in Daqinggou[J].Acta Phytoecologica Sinica,1999,23(5):475-479.
[15]邓永利,张峰,刘莹,等.万家寨引黄工程北干线沿线植被优势种群生态位[J].生态学杂志,2013,32(9):2263-2267.
DENG Y L,ZHANG F,LIU Y,et al.Niches of dominant populations in vegetation along the North Trunk of Wanjiazhai Yellow River Diversion Project[J].Chinese Journal of Ecology, 2013,32(9):2263-2267.
[16]高平,张贵平,吴琼,等.山西崦山自然保护区侧柏群落优势种群种间关系分析[J].植物研究,2010,30(6):731-736. GAO P,ZHANG G P,WU Q,et al.Analysis of interspecific
relationships among the dominant populations ofPlatycladus orientaliscommunities in Yanshan Nature Reserve in Shanxi[J]. Bulletin of Botanical Research,2010,30(6):731-736.
[17]方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
FANG J Y,WANG X P,SHEN Z H,et al.Methods and protocols for plant community inventory[J].Biodiversity Science, 2009,17(6):533-548.
[18]张峰.实验八:种群空间分布格局分析//傅必谦.生态学实验原理与方法[M].北京:科学出版社,2006:91-101.
ZHANG F.Experiment 8:analysis on population spatial pattern [M]∥FU B Q.Experiment principle and methods of ecology. Beijing:Science Press,2006:91-101.
[19]张桂萍,张峰,茹文明.山西绵山植被优势种群生态位研究[J].植物研究,2006,26(2):176-181.
ZHANG G P,ZHANG F,RU W M.Niche characteristics of dominant populations of vegetation in Mian Mountain,Shanxi[J]. Bulletin of Botanical Research,2006,26(2):176-181.
[20]毛空,张殷波,张峰.关帝山森林植被优势种群生态位[J].生态学杂志,2013,32(11):2920-2925.
MAO K,ZHANG Y B,ZHANG F.Niche characteristics of dominant species in forest vegetation in Guandi Mountain,Shanxi of China[J].Chinese Journal of Ecology,2013,32(11):2920-2925.
[21]彭少麟.鼎湖山森林群落植物优势种群生态位重叠的研究[J].热带亚热带森林生态系统研究,1990(6):19-27.
PENG S L.Niche overlap of dominant populations of forest communities in Dinghu Mountains,Guangdong[J].Researches on Forest Ecosystems in Tropics and Subtropics,1990(6):19-27.
(责任编辑 赵 勃责任编委 李景文)
Niche characteristics of dom inants in Platycladus orientalis forests in Yanshan Nature Reserve,Shanxi.
We studied the niche breadth and niche overlapping of the dominants inPlatycladus orientalisforests in Yanshan Nature Reserve,Shanxi Province by applying Shannon-Wiener index and niche special and general overlapping indices(Petraitis indices).The results indicated that the niche breadth ofP. orientaliswas the largest(3.56)andPinus bungeanathe second(3.12)among all dominant species. For the niche breadth of shrub species,Vitex negundovar.heterophyllawas the largest(3.43),Rosa xanthinaandLespedeza floribundawere tied for second(3.09),while for herbaceous species,Carex lanceolata(3.39)andArtemisia brachyloba(3.06)exceeded others.It was mainly because of their extensive adaptability to the environment.The relationship between niche breadth and distribution frequency of the species was positive(r=0.941,P<0.01),indicating that the lager the distribution area,the greater the ecological adaptability to the environment for a species,and thus a wider niche breadth could be.All dominant species were not extensively overlapping in the communities due to high environmental heterogeneity and mutual ecological complementation among species.A total of 1 099 species-pairs had niche overlapping,accounting for 87.22%of the total 1 260 species-pairs from 36 dominants.There was little niche overlapping betweenP.orientalisand other species,implying thatP. orientalisismore competitive and extensively adaptive than the other species.
Key words:Platycladus orientaliscommunity;dominant;niche breadth;niche overlapping;Yanshan Nature Reserve
中图分类号:S718.5
文献标识码:A
文章编号:1000-1522(2015)08-0024-07
收稿日期:2015-02-07
修回日期:2015-03-13
基金项目:山西省自然科学基金项目(2013011037-1)、科技部科技基础性工作专项(2011FY110300)。
DOI:10.13332/j.1000-1522.20150039